新規記事の投稿を行うことで、非表示にすることが可能です。
2023年02月12日
Alchemy of Actor Signal transductionシグナル伝達 05
Alchemy of Actor Signal transduction 05
④Gタンパク質共役型受容体(Gi)・・・アセチルコリン受容体(M2、M4)
三量体Gタンパク質にはGαの違いによって大きく分けてGs、Gi、Gqの3種類あり。
Gsはアデニル酸シクラーゼを活性化するタイプ、
Giはアデニル酸シクラーゼを抑制するタイプ、
GqはPLC(ホスホリパーゼC)を活性化するタイプ。
アセチルコリン受容体には、
イオンチャネル性の受容体と Gタンパク質共役型受容体の2種類あり。
ムスカリン性アセチルコリン受容体 (Gタンパク質共役型のアセチルコリン受容体のこと)、
M1~M5の5種類あり。
このうち、M2とM4はGiと共役しているGタンパク質共役型受容体(Gi)。
そのため、アセチルコリンが
7回膜貫通型のGタンパク質共役型受容体
ムスカリン性アセチルコリン受容体(M2あるいはM4)に結合すると、アデニル酸シクラーゼが抑制。
・ムスカリン性アセチルコリン受容体(M2とM4)はGiと共役しているGタンパク質共役型受容体(Gi)
・Gαiはアデニル酸シクラーゼを抑制し、細胞内のcAMP濃度を減少させる
⑤Gタンパク質共役型受容体(Gq)・・・アセチルコリン受容体(M1、M3、M5
Gタンパク質共役型のアセチルコリン受容体のうち、
M1とM3とM5はGqと共役しているGタンパク質共役型受容体(Gq)。
そのため、アセチルコリンが
7回膜貫通型のGタンパク質共役型受容体ムスカリン性アセチルコリン受容体(M1,M3.M5)に結合すると、
ホスホリパーゼC(PLC)を活性化、
ホスファチジルイノシトール4,5-ビスリン酸(PIP2)を分解、
ジアシルグリセロール(DG)とイノシトール3リン酸(IP3:イノシトール1,4,5-三リン酸)を産生。
ジアシルグリセロール(DG)とイノシトール3リン酸(IP3)はセカンドメッセンジャーとして働き、
ジアシルグリセロール(DG)は細胞膜上でプロテインキナーゼC(PKC)を活性化。一方、
イノシトール3リン酸(IP3)は小胞体からのCa2+の放出を促進、
プロテインキナーゼC(PKC)を活性化、
カルモジュリン/Ca2+結合タンパク質を介して
カルモジュリン依存性プロテインキナーゼ(CaMキナーゼ)も活性化。
・ムスカリン性アセチルコリン受容体(M1,M3,M5)は
Gqと共役しているGタンパク質共役型受容体(Gq)
・GαqはホスホリパーゼC(PLC)を活性化し、
ジアシルグリセロール(DG)とイノシトール3リン酸(IP3)を産生
と たのしい演劇の日々
④Gタンパク質共役型受容体(Gi)・・・アセチルコリン受容体(M2、M4)
三量体Gタンパク質にはGαの違いによって大きく分けてGs、Gi、Gqの3種類あり。
Gsはアデニル酸シクラーゼを活性化するタイプ、
Giはアデニル酸シクラーゼを抑制するタイプ、
GqはPLC(ホスホリパーゼC)を活性化するタイプ。
アセチルコリン受容体には、
イオンチャネル性の受容体と Gタンパク質共役型受容体の2種類あり。
ムスカリン性アセチルコリン受容体 (Gタンパク質共役型のアセチルコリン受容体のこと)、
M1~M5の5種類あり。
このうち、M2とM4はGiと共役しているGタンパク質共役型受容体(Gi)。
そのため、アセチルコリンが
7回膜貫通型のGタンパク質共役型受容体
ムスカリン性アセチルコリン受容体(M2あるいはM4)に結合すると、アデニル酸シクラーゼが抑制。
・ムスカリン性アセチルコリン受容体(M2とM4)はGiと共役しているGタンパク質共役型受容体(Gi)
・Gαiはアデニル酸シクラーゼを抑制し、細胞内のcAMP濃度を減少させる
⑤Gタンパク質共役型受容体(Gq)・・・アセチルコリン受容体(M1、M3、M5
Gタンパク質共役型のアセチルコリン受容体のうち、
M1とM3とM5はGqと共役しているGタンパク質共役型受容体(Gq)。
そのため、アセチルコリンが
7回膜貫通型のGタンパク質共役型受容体ムスカリン性アセチルコリン受容体(M1,M3.M5)に結合すると、
ホスホリパーゼC(PLC)を活性化、
ホスファチジルイノシトール4,5-ビスリン酸(PIP2)を分解、
ジアシルグリセロール(DG)とイノシトール3リン酸(IP3:イノシトール1,4,5-三リン酸)を産生。
ジアシルグリセロール(DG)とイノシトール3リン酸(IP3)はセカンドメッセンジャーとして働き、
ジアシルグリセロール(DG)は細胞膜上でプロテインキナーゼC(PKC)を活性化。一方、
イノシトール3リン酸(IP3)は小胞体からのCa2+の放出を促進、
プロテインキナーゼC(PKC)を活性化、
カルモジュリン/Ca2+結合タンパク質を介して
カルモジュリン依存性プロテインキナーゼ(CaMキナーゼ)も活性化。
・ムスカリン性アセチルコリン受容体(M1,M3,M5)は
Gqと共役しているGタンパク質共役型受容体(Gq)
・GαqはホスホリパーゼC(PLC)を活性化し、
ジアシルグリセロール(DG)とイノシトール3リン酸(IP3)を産生
と たのしい演劇の日々
【このカテゴリーの最新記事】
-
no image
-

-

-
no image
-
no image
2023年02月10日
Alchemy of Actor Signal transductionシグナル伝達 04
Alchemy of Actor Singlal trandsuction 04
②EGFやFGFなどの受容体・・・酵素共役型受容体(チロシンキナーゼ型受容体)
MAPK(MAPキナーゼ)のリン酸化の連鎖反応: MAPキナーゼカスケード
EGFやFGFなどの成長因子が受容体に結合すると、
自己リン酸化とアダプタータンパク質のリン酸化を介し、
GRB2とSOSの複合体が活性化される。
SOSは低分子量Gタンパク質であるRasに対するGEFとして作用、Rasを活性化。
活性化されたRasは、MAPKKK(MAPキナーゼキナーゼキナーゼ)であるRafをリン酸化して活性化。
活性化されたRafはMAPKK(MAPキナーゼキナーゼ)であるMEKをリン酸化して活性化。
そして、活性化されたMEKはMAPK(MAPキナーゼ)であるERKをリン酸化して活性化。
このようにして、Ras-Raf-MEK-ERKの経路(ERK経路)が活性化される。
MAPキナーゼカスケードには、ERK経路の他にもp38経路、JNK経路などがあり。
一方、EGFやFGFなどの受容体は
インスリン受容体と同じチロシンキナーゼ型の受容体なので、PI3K-Akt経路も活性化する。
③Gタンパク質共役型受容体(Gs)・・・グルカゴン受容体
グルカゴンが7回膜貫通型のGタンパク質共役型受容体であるグルカゴン受容体に結合すると、
α,β,γの三つのサブユニットから成るGタンパク質のαサブユニットからGDPが解離し、
GTPが結合するグアニンヌクレオチド交換反応が起こる。これによりGタンパク質は活性化。
その後、GTPが結合した活性型Gタンパク質はアデニル酸シクラーゼを活性化、
これがATPからcAMPへの合成を促進。
cAMPはセカンドメッセンジャーとしてPKA(プロテインキナーゼA)を活性化。
PKAとは、cAMPによって活性化されるプロテインキナーゼのこと。
7回膜貫通型Gタンパク質共役型受容体であるグルカゴン受容体に共役しているGαサブユニットは、
アデニル酸シクラーゼを活性化し細胞内cAMP濃度を上昇させるが、
このようにアデニル酸シクラーゼを活性化するタイプのGタンパク質Gsと呼ぶ。
・グルカゴン受容体はGsと共役しているGタンパク質共役型受容体(Gs)
・Gαsはアデニル酸シクラーゼを活性化し、細胞内のcAMP濃度を上昇
・cAMPはセカンドメッセンジャーとしてPKAを活性化しシグナル伝達を行う
と たのしい演劇の日々
②EGFやFGFなどの受容体・・・酵素共役型受容体(チロシンキナーゼ型受容体)
MAPK(MAPキナーゼ)のリン酸化の連鎖反応: MAPキナーゼカスケード
EGFやFGFなどの成長因子が受容体に結合すると、
自己リン酸化とアダプタータンパク質のリン酸化を介し、
GRB2とSOSの複合体が活性化される。
SOSは低分子量Gタンパク質であるRasに対するGEFとして作用、Rasを活性化。
活性化されたRasは、MAPKKK(MAPキナーゼキナーゼキナーゼ)であるRafをリン酸化して活性化。
活性化されたRafはMAPKK(MAPキナーゼキナーゼ)であるMEKをリン酸化して活性化。
そして、活性化されたMEKはMAPK(MAPキナーゼ)であるERKをリン酸化して活性化。
このようにして、Ras-Raf-MEK-ERKの経路(ERK経路)が活性化される。
MAPキナーゼカスケードには、ERK経路の他にもp38経路、JNK経路などがあり。
一方、EGFやFGFなどの受容体は
インスリン受容体と同じチロシンキナーゼ型の受容体なので、PI3K-Akt経路も活性化する。
③Gタンパク質共役型受容体(Gs)・・・グルカゴン受容体
グルカゴンが7回膜貫通型のGタンパク質共役型受容体であるグルカゴン受容体に結合すると、
α,β,γの三つのサブユニットから成るGタンパク質のαサブユニットからGDPが解離し、
GTPが結合するグアニンヌクレオチド交換反応が起こる。これによりGタンパク質は活性化。
その後、GTPが結合した活性型Gタンパク質はアデニル酸シクラーゼを活性化、
これがATPからcAMPへの合成を促進。
cAMPはセカンドメッセンジャーとしてPKA(プロテインキナーゼA)を活性化。
PKAとは、cAMPによって活性化されるプロテインキナーゼのこと。
7回膜貫通型Gタンパク質共役型受容体であるグルカゴン受容体に共役しているGαサブユニットは、
アデニル酸シクラーゼを活性化し細胞内cAMP濃度を上昇させるが、
このようにアデニル酸シクラーゼを活性化するタイプのGタンパク質Gsと呼ぶ。
・グルカゴン受容体はGsと共役しているGタンパク質共役型受容体(Gs)
・Gαsはアデニル酸シクラーゼを活性化し、細胞内のcAMP濃度を上昇
・cAMPはセカンドメッセンジャーとしてPKAを活性化しシグナル伝達を行う
と たのしい演劇の日々
2023年02月04日
Alchemy of Actor Singlal trandsuctionシグナル伝達 03
Alchemy of Actor Singlal trandsuction 03
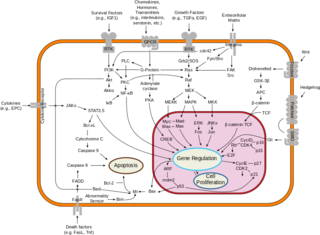
受容体を介した代表的なシグナル伝達の例
①インスリン受容体・・・酵素共役型受容体(チロシンキナーゼ型受容体)
「PI3K/Akt経路」インスリンの経路 その反応の酵素の名前から。
インスリンがインスリン受容体に結合すると、
インスリン受容体はインスリン受容体がもつチロシンキナーゼ活性により自己リン酸化。
これによってチロシンキナーゼ活性がさらに活性化され、
今度はアダプタータンパク質の一種IRS(insulin receptor substrate)をリン酸化。
次にリン酸化されたIRSはPI3キナーゼ(ホスファチジルイノシトールの3位をリン酸化する酵素)を活性化、細胞膜上のPIP2(ホスファチジルイノシトール2リン酸)を
PIP3(ホスファチジルイノシトール3リン酸)に変換。
このPIP3はドッキングサイトとしてプロテインキナーゼB(PKB、Akt)や
PDK1(ホスファチジルイノシトール依存性プロテインキナーゼ)を細胞膜に呼び寄せる。
その後、細胞膜に呼び寄せられたPKB(Akt)はPDK1によってリン酸化され、
活性型PKB(活性型Akt)に変換。
このようにして生じた活性型PKB(活性型Akt)は細胞膜を離れてさらに下流のタンパク質をリン酸化。
Aktの働きの一つにmTORC1(mechanistic target of rapamycin complex1)の活性化あり。
活性化されたAktはまず、
TSC2(Tuberous Sclerosis Complex 2:結節性硬化症複合体2)タンパク質をリン酸化して阻害。
TSC2は
TSC1やTBC1D7との3量体からなる複合体(TSC:TSC1-TSC2-TBC1D7)を形成しているが、
AktによりTSC2がリン酸化されることによって、リソソーム膜から解離。
TSCは、
mTORC1を活性化する低分子量Gタンパク質Rhebに対するGTPase活性化タンパク質
(GTPase- activating protein:GAP)として働くことで、
普段はmTORC1を阻害。
一方、活性化されたAktがTSC2をリン酸化してRhebの抑制が解除されると、
RhebによってmTORC1は活性化。
活性化されたmTORC1は、
標的タンパク質S6K(リン酸化されたS6Kは40Sリボソームタンパク質S6のリン酸化を介し翻訳を促進)や4EBP(リン酸化された4EBPは翻訳開始因子eIF4Eから解離し翻訳を促進)
をリン酸化することによって、タンパク質合成(翻訳)を促進。
・インスリン受容体はチロシンキナーゼ型受容体
・インスリンはPI3K-Akt経路を介してmTORC1を活性化
・mTORC1はS6Kや4EBPをリン酸化して翻訳を促進
と たのしい演劇の日々

受容体を介した代表的なシグナル伝達の例
①インスリン受容体・・・酵素共役型受容体(チロシンキナーゼ型受容体)
「PI3K/Akt経路」インスリンの経路 その反応の酵素の名前から。
インスリンがインスリン受容体に結合すると、
インスリン受容体はインスリン受容体がもつチロシンキナーゼ活性により自己リン酸化。
これによってチロシンキナーゼ活性がさらに活性化され、
今度はアダプタータンパク質の一種IRS(insulin receptor substrate)をリン酸化。
次にリン酸化されたIRSはPI3キナーゼ(ホスファチジルイノシトールの3位をリン酸化する酵素)を活性化、細胞膜上のPIP2(ホスファチジルイノシトール2リン酸)を
PIP3(ホスファチジルイノシトール3リン酸)に変換。
このPIP3はドッキングサイトとしてプロテインキナーゼB(PKB、Akt)や
PDK1(ホスファチジルイノシトール依存性プロテインキナーゼ)を細胞膜に呼び寄せる。
その後、細胞膜に呼び寄せられたPKB(Akt)はPDK1によってリン酸化され、
活性型PKB(活性型Akt)に変換。
このようにして生じた活性型PKB(活性型Akt)は細胞膜を離れてさらに下流のタンパク質をリン酸化。
Aktの働きの一つにmTORC1(mechanistic target of rapamycin complex1)の活性化あり。
活性化されたAktはまず、
TSC2(Tuberous Sclerosis Complex 2:結節性硬化症複合体2)タンパク質をリン酸化して阻害。
TSC2は
TSC1やTBC1D7との3量体からなる複合体(TSC:TSC1-TSC2-TBC1D7)を形成しているが、
AktによりTSC2がリン酸化されることによって、リソソーム膜から解離。
TSCは、
mTORC1を活性化する低分子量Gタンパク質Rhebに対するGTPase活性化タンパク質
(GTPase- activating protein:GAP)として働くことで、
普段はmTORC1を阻害。
一方、活性化されたAktがTSC2をリン酸化してRhebの抑制が解除されると、
RhebによってmTORC1は活性化。
活性化されたmTORC1は、
標的タンパク質S6K(リン酸化されたS6Kは40Sリボソームタンパク質S6のリン酸化を介し翻訳を促進)や4EBP(リン酸化された4EBPは翻訳開始因子eIF4Eから解離し翻訳を促進)
をリン酸化することによって、タンパク質合成(翻訳)を促進。
・インスリン受容体はチロシンキナーゼ型受容体
・インスリンはPI3K-Akt経路を介してmTORC1を活性化
・mTORC1はS6Kや4EBPをリン酸化して翻訳を促進
と たのしい演劇の日々
2023年02月03日
Alchemy of Actor Signal transductionシグナル伝達 02
Alchemy of Actor Signal transduction02
シグナル伝達の仕組みを理解するために、
どのようにしてシグナル分子が活性化されたり不活性化されるのかを探る。
シグナル伝達の様式、
・Gタンパク質guanine nucleotide-binding proteins, を介したシグナル伝達
・リン酸化phosphorylationを介したシグナル伝達
・セカンドメッセンジャーSecond messenger systemを介したシグナル伝達
【Gタンパク質】
とは、GTP結合タンパク質のこと。
GDPが結合している状態が不活性型で、GTPが結合している状態が活性型。
Gタンパク質は普段はGDPと結合した不活性型のGタンパク質として存在が、
刺激を受け、
Gタンパク質からGDP解離しGTP結合のグアニンヌクレオチド交換反応が起こり、
活性型のGタンパク質へ変換、シグナル伝達が行われる。
Gタンパク質は三量体Gタンパク質と低分子量Gタンパク質の2種類。
Gタンパク質共役型受容体(GPCR:G Protein-Coupled Receptor)、
この三量体Gタンパク質を介してシグナル伝達を行う、
【リン酸化を介したシグナル伝達】
多くのタンパク質は、さまざまな化学修飾を受けることによって
コンフォメーションや酵素活性などを変化させる。
タンパク質の化学修飾は、mRNAがタンパク質へと翻訳された後に行われるタンパク質の機能制御の機構で、特にタンパク質の翻訳後修飾と呼ぶ。
タンパク質の翻訳後修飾の代表的なものは、リン酸化
その他 アセチル化、メチル化、ユビキチン化、脂質付加、糖鎖付加などがあり。
タンパク質は、キナーゼ(リン酸化酵素)によってリン酸化され、
ホスファターゼ(脱リン酸化酵素)によって脱リン酸化される。
多くのタンパク質は、リン酸化や脱リン酸化を介して活性が調節される。
そのため、刺激によってキナーゼによるリン酸化
ホスファターゼによる脱リン酸化を受けることで、
タンパク質が活性化されシグナル伝達が行われる。
またリン酸化により活性化されるタンパク質も多いが、
中にはリン酸化により不活性されるタンパク質もあり。
タンパク質の翻訳後修飾の主な働き
・リン酸化・・・タンパク質の活性の制御など
・アセチル化やメチル化・・・タンパク質の活性の制御など
・ポリユビキチン化・・・プロテアソームによる分解の指標
・脂質付加・・・細胞内局在などを制御(細胞膜への埋め込み、小胞体膜への埋め込みなど)
・糖鎖付加・・・膜タンパク質に多く見られる
【セカンドメッセンジャー】
とは細胞膜で受容体が受け取ったシグナルを細胞内で中継する分子のこと。
ちなみに、受容体に結合するシグナル分子がファーストメッセンジャー。
セカンドメッセンジャー、
・cAMP
・ジアシルグリセロール(DG)とイノシトール3リン酸(IP3:イノシトール1,4,5-三リン酸)
・Ca2+
などがあり。
以下、これらの
セカンドメッセンジャーを産生するエフェクター分子と
セカンドメッセンジャーにより活性化される分子をまとめ。
・アデニル酸シクラーゼ→cAMP→プロテインキナーゼA(PKA)
・ホスホリパーゼC(PLC)→ジアシルグリセロール(DG)とイノシトール3リン酸(IP3)→
プロテインキナーゼC(PKC)やカルモジュリン依存性プロテインキナーゼ(CaMキナーゼ)
・イオンチャネル→Ca2+→カルモジュリン依存性プロテインキナーゼ(CaMキナーゼ)
と たのしい演劇の日々
シグナル伝達の仕組みを理解するために、
どのようにしてシグナル分子が活性化されたり不活性化されるのかを探る。
シグナル伝達の様式、
・Gタンパク質guanine nucleotide-binding proteins, を介したシグナル伝達
・リン酸化phosphorylationを介したシグナル伝達
・セカンドメッセンジャーSecond messenger systemを介したシグナル伝達
【Gタンパク質】
とは、GTP結合タンパク質のこと。
GDPが結合している状態が不活性型で、GTPが結合している状態が活性型。
Gタンパク質は普段はGDPと結合した不活性型のGタンパク質として存在が、
刺激を受け、
Gタンパク質からGDP解離しGTP結合のグアニンヌクレオチド交換反応が起こり、
活性型のGタンパク質へ変換、シグナル伝達が行われる。
Gタンパク質は三量体Gタンパク質と低分子量Gタンパク質の2種類。
Gタンパク質共役型受容体(GPCR:G Protein-Coupled Receptor)、
この三量体Gタンパク質を介してシグナル伝達を行う、
【リン酸化を介したシグナル伝達】
多くのタンパク質は、さまざまな化学修飾を受けることによって
コンフォメーションや酵素活性などを変化させる。
タンパク質の化学修飾は、mRNAがタンパク質へと翻訳された後に行われるタンパク質の機能制御の機構で、特にタンパク質の翻訳後修飾と呼ぶ。
タンパク質の翻訳後修飾の代表的なものは、リン酸化
その他 アセチル化、メチル化、ユビキチン化、脂質付加、糖鎖付加などがあり。
タンパク質は、キナーゼ(リン酸化酵素)によってリン酸化され、
ホスファターゼ(脱リン酸化酵素)によって脱リン酸化される。
多くのタンパク質は、リン酸化や脱リン酸化を介して活性が調節される。
そのため、刺激によってキナーゼによるリン酸化
ホスファターゼによる脱リン酸化を受けることで、
タンパク質が活性化されシグナル伝達が行われる。
またリン酸化により活性化されるタンパク質も多いが、
中にはリン酸化により不活性されるタンパク質もあり。
タンパク質の翻訳後修飾の主な働き
・リン酸化・・・タンパク質の活性の制御など
・アセチル化やメチル化・・・タンパク質の活性の制御など
・ポリユビキチン化・・・プロテアソームによる分解の指標
・脂質付加・・・細胞内局在などを制御(細胞膜への埋め込み、小胞体膜への埋め込みなど)
・糖鎖付加・・・膜タンパク質に多く見られる
【セカンドメッセンジャー】
とは細胞膜で受容体が受け取ったシグナルを細胞内で中継する分子のこと。
ちなみに、受容体に結合するシグナル分子がファーストメッセンジャー。
セカンドメッセンジャー、
・cAMP
・ジアシルグリセロール(DG)とイノシトール3リン酸(IP3:イノシトール1,4,5-三リン酸)
・Ca2+
などがあり。
以下、これらの
セカンドメッセンジャーを産生するエフェクター分子と
セカンドメッセンジャーにより活性化される分子をまとめ。
・アデニル酸シクラーゼ→cAMP→プロテインキナーゼA(PKA)
・ホスホリパーゼC(PLC)→ジアシルグリセロール(DG)とイノシトール3リン酸(IP3)→
プロテインキナーゼC(PKC)やカルモジュリン依存性プロテインキナーゼ(CaMキナーゼ)
・イオンチャネル→Ca2+→カルモジュリン依存性プロテインキナーゼ(CaMキナーゼ)
と たのしい演劇の日々
2023年02月02日
Alchemy of Actor Signal transductionシグナル伝達 01
Alchemy of Actor Signal transduction01
細胞がどのようにして細胞外のシグナル分子を認識するのか?
細胞外のシグナル分子は、リガンド(受容体に特異的に結合する物質)として、細胞膜あるいは細胞内に存在する受容体に結合し、細胞内シグナル伝達を引き起こす。
受容体を2つのタイプに分類。
①細胞膜受容体・・・Gタンパク質共役型受容体、酵素共役型受容体、イオンチャネル共役型受容体
②細胞内受容体・・・核内受容体
【細胞膜受容体】
『Gタンパク質共役型受容体GPCR』
水溶性のシグナル分子は、脂質二重層からなる細胞膜を通過することはできないので、
細胞膜に存在する受容体を介しシグナル伝達す。
Gタンパク質:グアニンヌクレオチド結合タンパク質。
は「GDPあるいはGTPが結合するタンパク質の総称」。
このGタンパク質はGDPが結合している状態が不活性型で、GTPが結合している状態が活性型になる。
そのため、Gタンパク質は、このGDPとGTPの結合状態を変化させることによって、
さまざまな刺激に応じて活性型と不活性型を行き来する。
このGタンパク質「三量体Gタンパク質」と「低分子量Gタンパク質」の2種類あり。
Gタンパク質共役型受容体は、この三量体Gタンパク質が結合している7回膜貫通型の受容体。
この7回膜貫通型の構造は、Gタンパク質共役型受容体の大きな特徴の一つ。
Gタンパク質共役型受容体にリガンドが結合するによって、Gタンパク質のαサブユニットからGDPが解離し、GTPが結合することで、細胞内シグナル伝達が進行。
この7回膜貫通型のGタンパク質共役型受容体には、アドレナリン受容体やグルカゴン受容体などがあり。
※現在市販されている薬の多くは、7回膜貫通型のGタンパク質共役型受容体をターゲット。
細胞内に存在し、受容体とは共役していない低分子量Gタンパク質には
GDPをGTPに交換する因子「グアニンヌクレオチド交換因子
(Guanine nucleotide Exchange Factor:GEF)」が結合し活性化す。
Gタンパク質自体は、弱いGTPase活性(GTPをGDPに加水分解する酵素活性)をもつが、
Gタンパク質自身では、活性化Gタンパク質を不活性型にすぐに戻すことはできん。
そこで、
細胞内にはGタンパク質がもつGTPase活性を活性化する
「GTPase活性化タンパク質(GTPase- activating protein:GAP)」因子が存在し
活性型のGタンパク質を不活性型へ戻す。
『酵素共役型受容体』
とは、自身が酵素活性をもつ受容体あるいは、酵素が直接結合している受容体のこと。
細胞外のシグナル分子がリガンドとして、受容体に結合すると細胞内の酵素が活性化し、
細胞内シグナル伝達が進行。
Exa,チロシンキナーゼ活性をもつインスリン受容体。
『イオンチャネル共役型受容体』
とは、リガンドが結合することでチャネルが開口し、細胞内外のイオンを通過させるタイプの受容体
【細胞内に存在する受容体(核内受容体)】
疎水性のシグナル分子は、
脂質二重層からなる細胞膜を通過することができるので、細胞内に取り込まれ、
細胞内に存在する受容体(核内受容体)を介しシグナル伝達。
※核内受容体とは、普段は細胞質に存在するが、
リガンド結合により核内へと移行、転写因子として標的遺伝子の発現を調節するタイプの受容体。
これら疎水性の物質 exa,ビタミンAやビタミンDなどの脂溶性ビタミン、ステロイドホルモンなどがあり。
と たのしい演劇の日々
細胞がどのようにして細胞外のシグナル分子を認識するのか?
細胞外のシグナル分子は、リガンド(受容体に特異的に結合する物質)として、細胞膜あるいは細胞内に存在する受容体に結合し、細胞内シグナル伝達を引き起こす。
受容体を2つのタイプに分類。
①細胞膜受容体・・・Gタンパク質共役型受容体、酵素共役型受容体、イオンチャネル共役型受容体
②細胞内受容体・・・核内受容体
【細胞膜受容体】
『Gタンパク質共役型受容体GPCR』
水溶性のシグナル分子は、脂質二重層からなる細胞膜を通過することはできないので、
細胞膜に存在する受容体を介しシグナル伝達す。
Gタンパク質:グアニンヌクレオチド結合タンパク質。
は「GDPあるいはGTPが結合するタンパク質の総称」。
このGタンパク質はGDPが結合している状態が不活性型で、GTPが結合している状態が活性型になる。
そのため、Gタンパク質は、このGDPとGTPの結合状態を変化させることによって、
さまざまな刺激に応じて活性型と不活性型を行き来する。
このGタンパク質「三量体Gタンパク質」と「低分子量Gタンパク質」の2種類あり。
Gタンパク質共役型受容体は、この三量体Gタンパク質が結合している7回膜貫通型の受容体。
この7回膜貫通型の構造は、Gタンパク質共役型受容体の大きな特徴の一つ。
Gタンパク質共役型受容体にリガンドが結合するによって、Gタンパク質のαサブユニットからGDPが解離し、GTPが結合することで、細胞内シグナル伝達が進行。
この7回膜貫通型のGタンパク質共役型受容体には、アドレナリン受容体やグルカゴン受容体などがあり。
※現在市販されている薬の多くは、7回膜貫通型のGタンパク質共役型受容体をターゲット。
細胞内に存在し、受容体とは共役していない低分子量Gタンパク質には
GDPをGTPに交換する因子「グアニンヌクレオチド交換因子
(Guanine nucleotide Exchange Factor:GEF)」が結合し活性化す。
Gタンパク質自体は、弱いGTPase活性(GTPをGDPに加水分解する酵素活性)をもつが、
Gタンパク質自身では、活性化Gタンパク質を不活性型にすぐに戻すことはできん。
そこで、
細胞内にはGタンパク質がもつGTPase活性を活性化する
「GTPase活性化タンパク質(GTPase- activating protein:GAP)」因子が存在し
活性型のGタンパク質を不活性型へ戻す。
『酵素共役型受容体』
とは、自身が酵素活性をもつ受容体あるいは、酵素が直接結合している受容体のこと。
細胞外のシグナル分子がリガンドとして、受容体に結合すると細胞内の酵素が活性化し、
細胞内シグナル伝達が進行。
Exa,チロシンキナーゼ活性をもつインスリン受容体。
『イオンチャネル共役型受容体』
とは、リガンドが結合することでチャネルが開口し、細胞内外のイオンを通過させるタイプの受容体
【細胞内に存在する受容体(核内受容体)】
疎水性のシグナル分子は、
脂質二重層からなる細胞膜を通過することができるので、細胞内に取り込まれ、
細胞内に存在する受容体(核内受容体)を介しシグナル伝達。
※核内受容体とは、普段は細胞質に存在するが、
リガンド結合により核内へと移行、転写因子として標的遺伝子の発現を調節するタイプの受容体。
これら疎水性の物質 exa,ビタミンAやビタミンDなどの脂溶性ビタミン、ステロイドホルモンなどがあり。
と たのしい演劇の日々
2023年01月31日
Alchemy of Actor Signal transductionシグナル伝達 00
Alchemy of Actor Signal transductionシグナル伝達 00
食事摂取で血糖値が上昇すれば、
インスリン(膵β細胞から分泌)は、標的細胞(骨格筋や脂肪組織など)に作用し糖取り込みが促進、
血糖値を低下させる。
これらの過程では、
まず「血糖値上昇」という刺激を受容し、
「インスリン」ホルモンの形で情報を伝達(情報を伝達する物質:シグナル分子)。
この情報が標的細胞に伝達され
さらに細胞内でシグナル伝達が連鎖反応(シグナルが急速に増幅していく様式:カスケード)、
最終的に「血糖値低下」という応答が起こる。
このとき、細胞が細胞外のシグナル分子を感知し、細胞内にその情報を伝えていく過程:シグナル伝達
細胞はさまざまなシグナル分子の情報を同時に受け取り、細胞内でさまざまなシグナル伝達が交差し合う。別々のシグナル伝達が互いに独立した経路をたどるだけでなく、
それぞれが互いに影響しあって作用する:シグナル伝達のクロストーク
シグナル伝達 の3つの段階。
「細胞間シグナル伝達」
「細胞におけるシグナル分子の受容(受容体による認識)」
「細胞内シグナル伝達」
【細胞間シグナル伝達】の5種類の様式。
①エンドクリン型(内分泌型):
細胞から分泌されたホルモンが血液を通って標的細胞まで運ばれるタイプのシグナル伝達の様式
②パラクリン型(傍分泌型):
細胞から分泌された物質が局所的に近隣した細胞に作用するタイプのシグナル伝達の様式
③オートクリン型(自己分泌型):
細胞から分泌された物質が自分自身に作用するタイプのシグナル伝達の様式。
分泌様式としては、パラクリン型(傍分泌型)と同じ
④細胞接触型:
接触しあった細胞同士がその細胞の表面に提示した物質を通じて情報を伝達するタイプのシグナル伝達の様式(分泌型ではない)
⑤神経型(シナプス型):
神経細胞のシナプスから神経伝達物質が放出されることによって、情報を迅速に伝達するタイプのシグナル伝達の様式
と たのしい演劇の日々
食事摂取で血糖値が上昇すれば、
インスリン(膵β細胞から分泌)は、標的細胞(骨格筋や脂肪組織など)に作用し糖取り込みが促進、
血糖値を低下させる。
これらの過程では、
まず「血糖値上昇」という刺激を受容し、
「インスリン」ホルモンの形で情報を伝達(情報を伝達する物質:シグナル分子)。
この情報が標的細胞に伝達され
さらに細胞内でシグナル伝達が連鎖反応(シグナルが急速に増幅していく様式:カスケード)、
最終的に「血糖値低下」という応答が起こる。
このとき、細胞が細胞外のシグナル分子を感知し、細胞内にその情報を伝えていく過程:シグナル伝達
細胞はさまざまなシグナル分子の情報を同時に受け取り、細胞内でさまざまなシグナル伝達が交差し合う。別々のシグナル伝達が互いに独立した経路をたどるだけでなく、
それぞれが互いに影響しあって作用する:シグナル伝達のクロストーク
シグナル伝達 の3つの段階。
「細胞間シグナル伝達」
「細胞におけるシグナル分子の受容(受容体による認識)」
「細胞内シグナル伝達」
【細胞間シグナル伝達】の5種類の様式。
①エンドクリン型(内分泌型):
細胞から分泌されたホルモンが血液を通って標的細胞まで運ばれるタイプのシグナル伝達の様式
②パラクリン型(傍分泌型):
細胞から分泌された物質が局所的に近隣した細胞に作用するタイプのシグナル伝達の様式
③オートクリン型(自己分泌型):
細胞から分泌された物質が自分自身に作用するタイプのシグナル伝達の様式。
分泌様式としては、パラクリン型(傍分泌型)と同じ
④細胞接触型:
接触しあった細胞同士がその細胞の表面に提示した物質を通じて情報を伝達するタイプのシグナル伝達の様式(分泌型ではない)
⑤神経型(シナプス型):
神経細胞のシナプスから神経伝達物質が放出されることによって、情報を迅速に伝達するタイプのシグナル伝達の様式
と たのしい演劇の日々
2023年01月16日
Alchemy of Actor epigenetics 16
Alchemy of Actor epigenetics 16
がん抑制遺伝子( tumor suppressor gene)は、
がん発生抑制機能を持つタンパク質(がん抑制タンパク質)をコードす(遺伝子p53、Rb、BRCA1)など。
2倍体の細胞において2つのがん抑制遺伝子両方が損傷することなどにより、
がん抑制タンパク質が作られなくなったり、
損傷遺伝子からの異常ながん抑制タンパク質が 正常がん抑制タンパク質の機能を阻害すると、
組織特異的にがん化が起きる。
現在十数以上のがん抑制遺伝子が知られており、組織特異的であることが多い。
p53の変異は大腸癌、乳癌など非組織特異的とみられる、
Rbの変異は網膜芽細胞腫、骨肉腫など、
BRCA1の変異は家族性乳がん、子宮がんなど、
MSH2の変異は大腸癌などに見られる。
これらがん抑制タンパク質の機能は
細胞周期チェックポイント制御、転写因子制御、転写、DNA修復など多岐にわたる。
癌でのDNAメチル化異常
がんでは ゲノム全体の低メチル化と CGI(CpG island)のメチル化 が特徴
LINEやAlu(SINE)などの繰返し配列はCPG部位に富み 正常細胞ではメチル化されている。
がん細胞ではこれら繰返し配列が低メチル化状態になり ゲノム全体の低メチル化として観察される。
低メチル化状態はゲノム不安定を招来し リンパ腫などの腫瘍発生を促進。
繰返し配列に加え 生理的にメチル化された特定のゲノム領域の低メチル化も起きる。
IGF2は 母親由来のアレルがメチル化により不活性化されている刷り込み遺伝子。
このIGF2の一定領域の脱メチル化により
本来不活性化されているはずの母親由来のアレルの転写が活性化される(Loss of Imprinting:LOI)。
大腸がんでは 増殖因子であるIGF2がLOIにより過剰に発現され 発がんを促進する。
正常細胞では 殆どのCGIは脱メチル化状態に保たれている。
がん細胞では 一部のCGIが異常にメチル化されている。
CGIのメチル化は 突然変異 染色体欠失同様に 遺伝子不活性化の原因となる。
p16, VHL, BRCA1, CDH1など
重要ながん制御遺伝子のメチル化による不活性化が 様々な癌で高頻度に見られる。
がん組織のヒストン
各ヒストンタンパク質のバリアント(アミノ酸配列が異なる変異体)が入れ替わることで、
クロマチン構造を変え、特異的な核内プロセスを制御す。
H2AファミリーのバリアントH2A.Xは、
DNAのダメージを監視し、DNA修復タンパク質のリクルートを促進し、
ゲノムの保全に働くバリアントH2A.Zは、遺伝子の活性抑制の双方で重要な役割を持つ。
高レベルのH2A.Z発現は、多くのがんで広範に検出され、細胞増殖とゲノムの不安定性とに関連。
がんに特異的なヒストンの化学的修飾。
がん抑制遺伝子プロモーターのCpGアイランドDNAメチル化は、
ヒストン脱アセチル化酵素 (HDAC) をリクルートすることで
がん抑制遺伝子の発現を抑制し、がんの発生の一因。
と たのしい演劇の日々
がん抑制遺伝子( tumor suppressor gene)は、
がん発生抑制機能を持つタンパク質(がん抑制タンパク質)をコードす(遺伝子p53、Rb、BRCA1)など。
2倍体の細胞において2つのがん抑制遺伝子両方が損傷することなどにより、
がん抑制タンパク質が作られなくなったり、
損傷遺伝子からの異常ながん抑制タンパク質が 正常がん抑制タンパク質の機能を阻害すると、
組織特異的にがん化が起きる。
現在十数以上のがん抑制遺伝子が知られており、組織特異的であることが多い。
p53の変異は大腸癌、乳癌など非組織特異的とみられる、
Rbの変異は網膜芽細胞腫、骨肉腫など、
BRCA1の変異は家族性乳がん、子宮がんなど、
MSH2の変異は大腸癌などに見られる。
これらがん抑制タンパク質の機能は
細胞周期チェックポイント制御、転写因子制御、転写、DNA修復など多岐にわたる。
癌でのDNAメチル化異常
がんでは ゲノム全体の低メチル化と CGI(CpG island)のメチル化 が特徴
LINEやAlu(SINE)などの繰返し配列はCPG部位に富み 正常細胞ではメチル化されている。
がん細胞ではこれら繰返し配列が低メチル化状態になり ゲノム全体の低メチル化として観察される。
低メチル化状態はゲノム不安定を招来し リンパ腫などの腫瘍発生を促進。
繰返し配列に加え 生理的にメチル化された特定のゲノム領域の低メチル化も起きる。
IGF2は 母親由来のアレルがメチル化により不活性化されている刷り込み遺伝子。
このIGF2の一定領域の脱メチル化により
本来不活性化されているはずの母親由来のアレルの転写が活性化される(Loss of Imprinting:LOI)。
大腸がんでは 増殖因子であるIGF2がLOIにより過剰に発現され 発がんを促進する。
正常細胞では 殆どのCGIは脱メチル化状態に保たれている。
がん細胞では 一部のCGIが異常にメチル化されている。
CGIのメチル化は 突然変異 染色体欠失同様に 遺伝子不活性化の原因となる。
p16, VHL, BRCA1, CDH1など
重要ながん制御遺伝子のメチル化による不活性化が 様々な癌で高頻度に見られる。
がん組織のヒストン
各ヒストンタンパク質のバリアント(アミノ酸配列が異なる変異体)が入れ替わることで、
クロマチン構造を変え、特異的な核内プロセスを制御す。
H2AファミリーのバリアントH2A.Xは、
DNAのダメージを監視し、DNA修復タンパク質のリクルートを促進し、
ゲノムの保全に働くバリアントH2A.Zは、遺伝子の活性抑制の双方で重要な役割を持つ。
高レベルのH2A.Z発現は、多くのがんで広範に検出され、細胞増殖とゲノムの不安定性とに関連。
がんに特異的なヒストンの化学的修飾。
がん抑制遺伝子プロモーターのCpGアイランドDNAメチル化は、
ヒストン脱アセチル化酵素 (HDAC) をリクルートすることで
がん抑制遺伝子の発現を抑制し、がんの発生の一因。
と たのしい演劇の日々
2023年01月09日
Alchemy of Actor epigenetics 15
Alchemy of Actor epigenetics 15
Paramutation・パラ変異
「動物の行動は、その行動を行う特定の動物の体内にその遺伝子が存在するかどうかに関わらず、
その行動のための遺伝子の生存を最大化する傾向がある。」
Extended Phenotype (1982 by Richard Dawkins (1941- evolutionary biologist),
パラミューテーション:
相同DNA配列が減数分裂的な遺伝的発現状態を確立するためにトランス状態で情報伝達する過程 ,
その機構は未だ完全には解明されていない。
パラミューテーションは特異的なクロマチン状態の確立と 遺伝的伝達の基礎になる かも。,
植物(Arabodpsis thalianaやトウモロコシ)と酵母(Saccharomyces pombe)での
RNA依存転写サイレンシングに関与する転写/
非コード性タンデム反復配列と蛋白質はパラミューテーションにとって必要 だが,
遺伝的サイレンシングに介入する特異的分子類は未決。
遺伝法則に反する遺伝パターン、
1950年代トウモロコシで発見、それ以来植物や菌類、マウスでも認められる。
メンデルの法則はほとんどの場合に当てはまるが、各対立遺伝子が独立して分離す。
表現型に現れないほうの対立遺伝子が、実際に発現する方の対立遺伝子を「変異」させ、
発現しないようにしてしまう相互作用。
トウモロコシの研究で、パラミューテーションはRNAによるものであることが示された。
トランスポゾンのサイレンシングに関係するクロマチン状態の安定性には、
RNA依存性RNAポリメラーゼをコードするmop1遺伝子が必要。
1対の対立遺伝子のそれぞれが分離して配偶子に振り分けられる とするメンデルの法則は、
たいていの場合に当てはまる。しかし、
1個の遺伝子座にある2個の対立遺伝子の相互作用であるパラミューテーションが生じると、
片方の対立遺伝子に遺伝性の変化が起こる。このような非メンデル遺伝が動物でも見つかった。
マウスのKit遺伝子研究、ヌル変異体との交雑で野生型の表現型の出現が抑えられる。
kit遺伝子:4番染色体長腕(4q12)に座位する遺伝子
このエピジェネティックな(DNAに依存しない)変異の仕組みは、
配偶子と接合子の間でのRNAの受け渡しが関係。
RNAはこの現象の立役者の1人で、遺伝情報の貯蔵庫、調節性のマイクロRNAとして、
さまざまなエピジェネティックな遺伝にかかわっている。
と たのしい演劇の日々
Paramutation・パラ変異
「動物の行動は、その行動を行う特定の動物の体内にその遺伝子が存在するかどうかに関わらず、
その行動のための遺伝子の生存を最大化する傾向がある。」
Extended Phenotype (1982 by Richard Dawkins (1941- evolutionary biologist),
パラミューテーション:
相同DNA配列が減数分裂的な遺伝的発現状態を確立するためにトランス状態で情報伝達する過程 ,
その機構は未だ完全には解明されていない。
パラミューテーションは特異的なクロマチン状態の確立と 遺伝的伝達の基礎になる かも。,
植物(Arabodpsis thalianaやトウモロコシ)と酵母(Saccharomyces pombe)での
RNA依存転写サイレンシングに関与する転写/
非コード性タンデム反復配列と蛋白質はパラミューテーションにとって必要 だが,
遺伝的サイレンシングに介入する特異的分子類は未決。
遺伝法則に反する遺伝パターン、
1950年代トウモロコシで発見、それ以来植物や菌類、マウスでも認められる。
メンデルの法則はほとんどの場合に当てはまるが、各対立遺伝子が独立して分離す。
表現型に現れないほうの対立遺伝子が、実際に発現する方の対立遺伝子を「変異」させ、
発現しないようにしてしまう相互作用。
トウモロコシの研究で、パラミューテーションはRNAによるものであることが示された。
トランスポゾンのサイレンシングに関係するクロマチン状態の安定性には、
RNA依存性RNAポリメラーゼをコードするmop1遺伝子が必要。
1対の対立遺伝子のそれぞれが分離して配偶子に振り分けられる とするメンデルの法則は、
たいていの場合に当てはまる。しかし、
1個の遺伝子座にある2個の対立遺伝子の相互作用であるパラミューテーションが生じると、
片方の対立遺伝子に遺伝性の変化が起こる。このような非メンデル遺伝が動物でも見つかった。
マウスのKit遺伝子研究、ヌル変異体との交雑で野生型の表現型の出現が抑えられる。
kit遺伝子:4番染色体長腕(4q12)に座位する遺伝子
このエピジェネティックな(DNAに依存しない)変異の仕組みは、
配偶子と接合子の間でのRNAの受け渡しが関係。
RNAはこの現象の立役者の1人で、遺伝情報の貯蔵庫、調節性のマイクロRNAとして、
さまざまなエピジェネティックな遺伝にかかわっている。
と たのしい演劇の日々

2022年12月20日
Alchemy of Actor epigenetics 14
Alchemy of Actor epigenetics 14
プリオンprion
は感染可能なタンパク質の形態。タンパク質からなる感染性因子のこと
ミスフォールド(誤って折りたたまれたタンパク質)状態のタンパク質が
その構造を正常の構造のタンパク質に伝えることによって伝播す。
一部のタンパク質は、複数の立体構造をとるように変化でき、その一例としてプリオンがある。
プリオンは、
同じアミノ酸配列のタンパク質を自然状態から感染性立体構造へ触媒的に変換するタンパク質と定義
ゲノムを変更せず表し 現型の変化を誘導することができるエピジェネティックな媒介物と見る。
プリオンは他の感染性因子と異なり、DNAやRNAといった核酸は含まれていない。
狂牛病やクロイツフェルト・ヤコブ病などの伝達性海綿状脳症の原因となり、
これらの病気はプリオン病と呼ばれる。
脳などの神経組織の構造に影響を及ぼす極めて進行が速い疾患として知られており、
治療法が確立していない致死性の疾患。
哺乳類においてプリオンとしてふるまい、狂牛病などのプリオン病の原因となるのはPrP。
PrPは、ヒトで253個、マウスで254個のアミノ酸からなるタンパク質、
そのアミノ酸配列は高度に保存される。
PrPは健康なヒトや動物でも発現しているタンパク質で、
脳、心臓、肝臓など多くの組織、臓器において発現、特に脳、神経細胞において高く発現。
同一のアミノ酸配列でありながら、
正常プリオンタンパク質と異常プリオンタンパク質の二つの異なる高次構造をとる、
異常プリオンタンパク質(scrapie PrP:PrPSc)
PrPCが構造変化を起こしたプリオン病に特異的に検出される。
PrP遺伝子はヒトにおいては第20番染色体上に存在し、2つのエクソンからなる。
PrPScは、PrPCと比べてβシート構造に富んだ構造
PrPCが界面活性剤に可溶性、プロテアーゼKなどのタンパク質分解酵素によって容易に分解されるのに対し、PrPScは、界面活性剤に難溶性、タンパク質分解酵素にも抵抗性を示す。
PrPScの凝集体はアミロイド線維とよばれる構造、
PrPのアミロイド線維はPrP単量体が結合する鋳型として働くことができ、
PrPの単量体がPrPのアミロイド線維にとりこまれることによってPrPのアミロイドは伸長する。
毒性・感染力の強いPrPScはアミロイドよりもむしろオリゴマー。
( oligome:)比較的少数のモノマーが結合した重合体のこと
有限個(10~100個)のモノマーが結合した比較的分子量が低い重合体を指す。
オリゴマーに対してポリマーは非常に多数(数100個以上)のモノマーが結合した状態のこと
ミスフォールドしたPrPが健康な個体に感染すると、
健康な個体に存在していた正常な構造のPrPがミスフォールドしたPrPへの構造変換が起きる。
ミスフォールドしたPrPは他のPrPの構造変換を引き起こす鋳型としてふるまい、
可溶性の正常型タンパク質がアミロイドに重合していくことによって、
構造変化がおこり異常型タンパク質の構造へと変化する。
アミロイドは物理的にも化学的にも非常に安定な構造であり、
このことがプリオン病の封じ込めを困難にしている。
と たのしい演劇の日々
プリオンprion
は感染可能なタンパク質の形態。タンパク質からなる感染性因子のこと
ミスフォールド(誤って折りたたまれたタンパク質)状態のタンパク質が
その構造を正常の構造のタンパク質に伝えることによって伝播す。
一部のタンパク質は、複数の立体構造をとるように変化でき、その一例としてプリオンがある。
プリオンは、
同じアミノ酸配列のタンパク質を自然状態から感染性立体構造へ触媒的に変換するタンパク質と定義
ゲノムを変更せず表し 現型の変化を誘導することができるエピジェネティックな媒介物と見る。
プリオンは他の感染性因子と異なり、DNAやRNAといった核酸は含まれていない。
狂牛病やクロイツフェルト・ヤコブ病などの伝達性海綿状脳症の原因となり、
これらの病気はプリオン病と呼ばれる。
脳などの神経組織の構造に影響を及ぼす極めて進行が速い疾患として知られており、
治療法が確立していない致死性の疾患。
哺乳類においてプリオンとしてふるまい、狂牛病などのプリオン病の原因となるのはPrP。
PrPは、ヒトで253個、マウスで254個のアミノ酸からなるタンパク質、
そのアミノ酸配列は高度に保存される。
PrPは健康なヒトや動物でも発現しているタンパク質で、
脳、心臓、肝臓など多くの組織、臓器において発現、特に脳、神経細胞において高く発現。
同一のアミノ酸配列でありながら、
正常プリオンタンパク質と異常プリオンタンパク質の二つの異なる高次構造をとる、
異常プリオンタンパク質(scrapie PrP:PrPSc)
PrPCが構造変化を起こしたプリオン病に特異的に検出される。
PrP遺伝子はヒトにおいては第20番染色体上に存在し、2つのエクソンからなる。
PrPScは、PrPCと比べてβシート構造に富んだ構造
PrPCが界面活性剤に可溶性、プロテアーゼKなどのタンパク質分解酵素によって容易に分解されるのに対し、PrPScは、界面活性剤に難溶性、タンパク質分解酵素にも抵抗性を示す。
PrPScの凝集体はアミロイド線維とよばれる構造、
PrPのアミロイド線維はPrP単量体が結合する鋳型として働くことができ、
PrPの単量体がPrPのアミロイド線維にとりこまれることによってPrPのアミロイドは伸長する。
毒性・感染力の強いPrPScはアミロイドよりもむしろオリゴマー。
( oligome:)比較的少数のモノマーが結合した重合体のこと
有限個(10~100個)のモノマーが結合した比較的分子量が低い重合体を指す。
オリゴマーに対してポリマーは非常に多数(数100個以上)のモノマーが結合した状態のこと
ミスフォールドしたPrPが健康な個体に感染すると、
健康な個体に存在していた正常な構造のPrPがミスフォールドしたPrPへの構造変換が起きる。
ミスフォールドしたPrPは他のPrPの構造変換を引き起こす鋳型としてふるまい、
可溶性の正常型タンパク質がアミロイドに重合していくことによって、
構造変化がおこり異常型タンパク質の構造へと変化する。
アミロイドは物理的にも化学的にも非常に安定な構造であり、
このことがプリオン病の封じ込めを困難にしている。
と たのしい演劇の日々