2019”N03Œژ04“ْ
DNA‚جگL’·•ûŒü‚ح5'پ¨3'•ûŒü‚ة‚ب‚é——R‚ًچ\‘¢ƒŒƒxƒ‹‚إ—‰ً‚µ‚و‚¤
چ،‰ٌ‚ح•ھژqگ¶•¨ٹw‚ج“à—e‚إ‚·پB
•ھژqگ¶•¨ٹw‚ھ•×‹‚µ‚ؤ‚¢‚é‚ئ•K‚¸ڈo‚ؤ‚‚éDNAچ½‚جگL’·•ûŒü‚ة‚آ‚¢‚ؤ‚جچl‚¦•û‚إ‚·پB
‚ـ‚¸پAٹî‘b‚ج“à—e‚ئ‚µ‚ؤDNA‚ئ‚¢‚¤‚ج‚ح‹t•ûŒüگ«‚ج2–{چ½ƒkƒNƒŒƒIƒ`ƒhچ\‘¢‚ًژو‚è‚ـ‚·‚ثپB
ƒCƒپپ[ƒW‚إ‚¢‚¤‚ئˆب‰؛‚ج‚و‚¤‚بچ\‘¢‚إ‚·پB
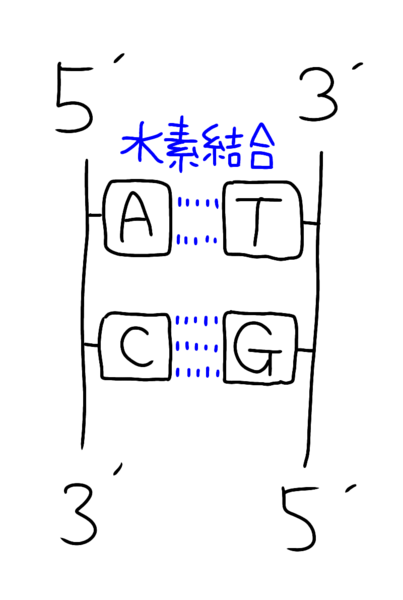
•ذ•û‚جچ½‚ھ3'پ¨5'•ûŒü‚¾‚ئپA‚à‚¤ˆê•û‚ح5'پ¨3'•ûŒü‚ة‚ب‚ء‚ؤ‚¢‚ـ‚·پiپپ‚±‚ê‚ھ‹t•ûŒüگ«پjپB
‚ب‚¨پAƒAƒfƒjƒ“پiAپj‚حƒ`ƒ~ƒ“پiTپj‚ئ‘خ‚ئ‚ب‚èگ…‘fŒ‹چ‡‚ح2–{
ƒVƒgƒVƒ“پiCپj‚حƒOƒAƒjƒ“پiGپj‚ئ‘خ‚ئ‚ب‚èپAگ…‘fŒ‹چ‡‚ح3–{‚إ‚·
پiپ¦CG‚حƒRƒ“ƒsƒ…پ[ƒ^پ[ƒOƒ‰ƒtƒBƒbƒN‚إ3D‚جƒCƒپپ[ƒW‚إ3–{‚ئٹo‚¦‚éپj
‚±‚±‚ـ‚إ‚ح–â‘è‚ب‚¢‚ئژv‚¤‚ج‚إ‚·‚ھپA
ˆّ‚ء‚©‚©‚ء‚ؤ‚µ‚ـ‚¤گl‚ھ‘‚¦‚é‚ج‚ھ
پu•،گ»پE“]ژتپE–|–َپv‚جچغ‚جپA•ûŒüگ«‚جکb‚ة‚ب‚ء‚½چغ‚إ‚·پB
پE•،گ»‚حDNA‚©‚çDNA‚ًچ‡گ¬‚·‚锽‰
پE“]ژت‚حDNA‚©‚çRNA‚ًچ‡گ¬‚·‚锽‰
پE–|–َ‚حmRNA‚©‚çƒ^ƒ“ƒpƒNژ؟‚ًچ‡گ¬‚·‚锽‰‚ًژw‚µ‚ـ‚·‚ھ
‚ـ‚ئ‚ك‚é‚ئˆب‰؛‚ج‚و‚¤‚ة‚ب‚è‚ـ‚·
ڈم‚ج•\‚ج“à—e‚ًٹo‚¦‚éچغ‚ةڈd—v‚ة‚ب‚ء‚ؤ‚‚é•”•ھ‚ًگشگF‚إژ¦‚µ‚ـ‚µ‚½پB
‹ة’[‚بکbپAڈم‚ج•\‚ج“à—e‚ًٹo‚¦‚邽‚ك‚ة‚حگشگF‚ج•”•ھ‚¾‚¯ٹo‚¦‚ê‚خ‚و‚¢‚ج‚إ‚·پB
‚آ‚ـ‚èپuƒkƒNƒŒƒIƒ`ƒh‚جگL’·•ûŒü‚ح5'پ¨3'•ûŒü‚ةŒہ’肳‚ê‚ؤ‚¢‚éپv
ƒkƒNƒŒƒIƒ`ƒh‚حپuƒSƒ~پi5'پ¨3'پj‚ج•ûŒü‚ة‚µ‚©گL‚خ‚¹‚ب‚¢پv‚ئ‚¢‚¤“à—e‚ھچإ‚àڈd—v‚بƒ|ƒCƒ“ƒg‚ة‚ب‚è‚ـ‚·پB
‚±‚ê‚ًٹغˆأ‹L‚·‚é‚ج‚إ‚ح‚ب‚پAچ\‘¢ƒŒƒxƒ‹‚إ—‰ً‚µ‚ـ‚µ‚ه‚¤پB
‰؛‚جƒCƒ‰ƒXƒg‚حƒkƒNƒŒƒIƒ`ƒh‚ھDNAƒ|ƒٹƒپƒ‰پ[ƒ[‚ب‚ا‚إگL’·‚µ‚ؤ‚¢‚‚ج‚ً–حژ®‰»‚µ‚½‚à‚ج‚ة‚ب‚è‚ـ‚·پB
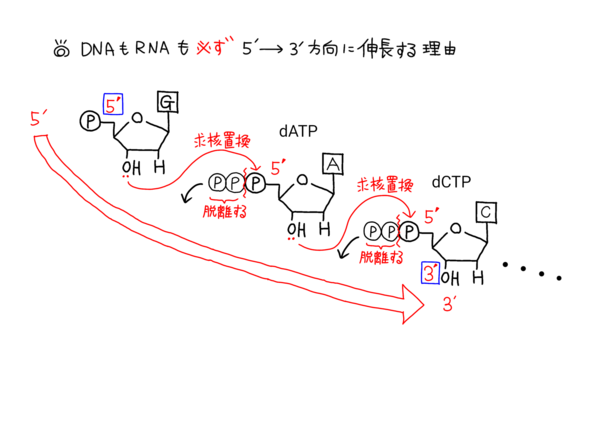
‚±‚جƒCƒ‰ƒXƒg‚ً‚و‚Œ©‚ؤ‚à‚炦‚خپADNAپi‚âRNA‚ئ‚¢‚ء‚½ƒkƒNƒŒƒIƒ`ƒhپj‚ھ5'پ¨3'•ûŒü‚ة‚µ‚©گL’·‚µ‚ب‚¢——R‚ھ‚و‚•ھ‚©‚é‚©‚ئژv‚¢‚ـ‚·پB
‚»‚µ‚ؤپA–`“ھ‚ةگ\‚µڈم‚°‚½‚ئ‚¨‚èپADNAپiƒkƒNƒŒƒIƒ`ƒhپj‚ح2–{چ½‚ًŒ`گ¬‚·‚éچغ‚ة‚ح‹t•ûŒüگ«‚ً‚ئ‚邽‚كپA’’Œ^پi“ا‚فژو‚ç‚ê‚鑤‚جچ½پj‚©‚ç‚جژ‹“_‚إ‚ح•،گ»‚ئ“]ژت‚ة‚¨‚¢‚ؤپA’’Œ^‚حپu3'پ¨5'پv•ûŒü‚ة“ا‚فژو‚ç‚ê‚ؤ‚¢‚‚±‚ئ‚ة‚ب‚éپB
‚±‚±‚ـ‚إ‚ج“à—e‚إپA‚±‚ج•\‚ج•،گ»پE“]ژت‚جچs‚ة‚آ‚¢‚ؤ‚حگà–¾‚ھ‚إ‚«‚ـ‚µ‚½پB
‚»‚ê‚إ‚حپA–|–َ‚ة‚آ‚¢‚ؤ‚حˆê‘ج‚ا‚ج‚و‚¤‚ةچl‚¦‚é‚ׂ«‚ب‚ج‚إ‚µ‚ه‚¤‚©پB
‚±‚±‚إ‘هژ–‚ب‚ج‚حپAپu–|–َپپƒkƒNƒŒƒIƒ`ƒh‚ًچى‚锽‰‚إ‚ح‚ب‚¢پv‚ئ‚¢‚¤‚±‚ئ‚إ‚·پB
پw–|–َپx‚ئ‚حmRNA‚ً‚à‚ئ‚ةƒ^ƒ“ƒpƒNژ؟‚ًچى‚锽‰‚ب‚ج‚إپA
’’Œ^‚ئ‚ب‚émRNA‚ح—ک_ڈمپA5'پ¨3'•ûŒü‚ة“ا‚فژو‚邱‚ئ‚ة‰½‚çژxڈل‚ح‚ب‚¢‚ج‚إ‚·پB
پiگ®—‚µ‚ؤ‚ظ‚µ‚¢‚ج‚حپAپw•،گ»پx‚ئپw–|–َپx‚حپA‚»‚ꂼ‚êDNA‚âRNA‚ئ‚¢‚¤ƒkƒNƒŒƒIƒ`ƒh‚ًچ‡گ¬‚·‚锽‰‚إ‚ ‚èپA
پ@ƒkƒNƒŒƒIƒ`ƒh‚جگL’·پiچ‡گ¬پj•ûŒü‚ة‚ح5'پ¨3'‚ج‚ف‚ئ‚¢‚¤”›‚è‚ھ‚ ‚èپA•K‘R“I‚ة’’Œ^‚ح3'پ¨5'‚ة“ا‚فژو‚ç‚ê‚é•K—v‚ھ‚ ‚ء‚½‚±‚ئپB–|–َ‚حƒ^ƒ“ƒpƒNژ؟‚ئ‚¢‚¤ƒ|ƒٹƒyƒvƒ`ƒh‚ًچى‚锽‰‚إ‚ ‚èپAN––’[پAC––’[‚ئ‚¢‚¤ٹT”O‚ح‚ ‚é‚ھپA3'‚ئ‚©5'‚ب‚ا‚جٹT”O‚ح•K—v‚ب‚¢‚ئ‚¢‚¤“_‚إ‚ ‚éپj
پw“]ژتپx‚جچغ‚ةmRNA‚ًچ‡گ¬پimRNA‚جƒvƒچƒZƒVƒ“ƒO‚ًپj‚·‚éچغپA
mRNA‚ج5'––’[‚ة‚ح7-ƒپƒ`ƒ‹ƒOƒAƒmƒVƒ“‚ھ•t‰ء‚³‚êپicapچ\‘¢‚ئŒؤ‚شپjپA3'––’[‚ة‚حƒ|ƒٹپiAپjƒeƒCƒ‹‚ئŒؤ‚خ‚ê‚éƒAƒfƒjƒ‹ژ_‚ھ150پ`200Œآ‚ظ‚اکAŒ‹‚µ‚½چ\‘¢‚ھ•t‰ء‚³‚ê‚éپB
‚»‚µ‚ؤmRNA‚حپA5'––’[‚جcapچ\‘¢‚©‚çƒٹƒ{ƒ\پ[ƒ€‚جڈ¬ƒTƒuƒ†ƒjƒbƒgپiگ^ٹjگ¶•¨‚إ‚ح40SپAŒ´ٹjگ¶•¨‚إ‚ح30SƒTƒuƒ†ƒjƒbƒgپj
‚ة‚و‚è“ا‚فژو‚ç‚ê‚ؤ3'––’[‚جƒ|ƒٹپiAپjƒeƒCƒ‹‚ج‚ظ‚¤‚ةŒü‚©‚ء‚ؤ“ا‚فژو‚ç‚ê‚ؤ‚¢‚پB
پ¦5'––’[‚جƒLƒƒƒbƒvچ\‘¢‚ج‹ك‚‚ة‘¶چف‚·‚éٹJژnƒRƒhƒ“‚©‚ç“ا‚فژو‚è‚ًƒXƒ^پ[ƒg‚µ‚ؤپA3'––’[‚جƒ|ƒٹپiAپjƒeƒCƒ‹‘¤‚ةڈIژ~ƒRƒhƒ“‚ھ‘¶چف‚·‚邽‚كپAپumRNA‚حƒLƒƒƒbƒv‚ً”ي‚ء‚ؤ‚¢‚é“ھپi5'پj‚©‚ç“ا‚فژو‚è‚ًٹJژn‚·‚éپv‚ئٹo‚¦‚é
‚µ‚½‚ھ‚ء‚ؤپAŒ‹ک_‚ئ‚µ‚ؤ–|–َ‚ة‚¨‚¢‚ؤ‚ح
’’Œ^mRNA‚ح5'پ¨3'•ûŒü‚ة“ا‚فژو‚ç‚ê‚ؤپA
ƒ^ƒ“ƒpƒNژ؟‚حN––’[پ¨C––’[•ûŒü‚ةگL’·‚ھگi‚ٌ‚إ‚¢‚‚±‚ئ‚ئ‚ب‚éپB
•ھژqگ¶•¨ٹw‚ھ•×‹‚µ‚ؤ‚¢‚é‚ئ•K‚¸ڈo‚ؤ‚‚éDNAچ½‚جگL’·•ûŒü‚ة‚آ‚¢‚ؤ‚جچl‚¦•û‚إ‚·پB
‚ـ‚¸پAٹî‘b‚ج“à—e‚ئ‚µ‚ؤDNA‚ئ‚¢‚¤‚ج‚ح‹t•ûŒüگ«‚ج2–{چ½ƒkƒNƒŒƒIƒ`ƒhچ\‘¢‚ًژو‚è‚ـ‚·‚ثپB
ƒCƒپپ[ƒW‚إ‚¢‚¤‚ئˆب‰؛‚ج‚و‚¤‚بچ\‘¢‚إ‚·پB
DNA‚ج‹t•ûŒüگ«
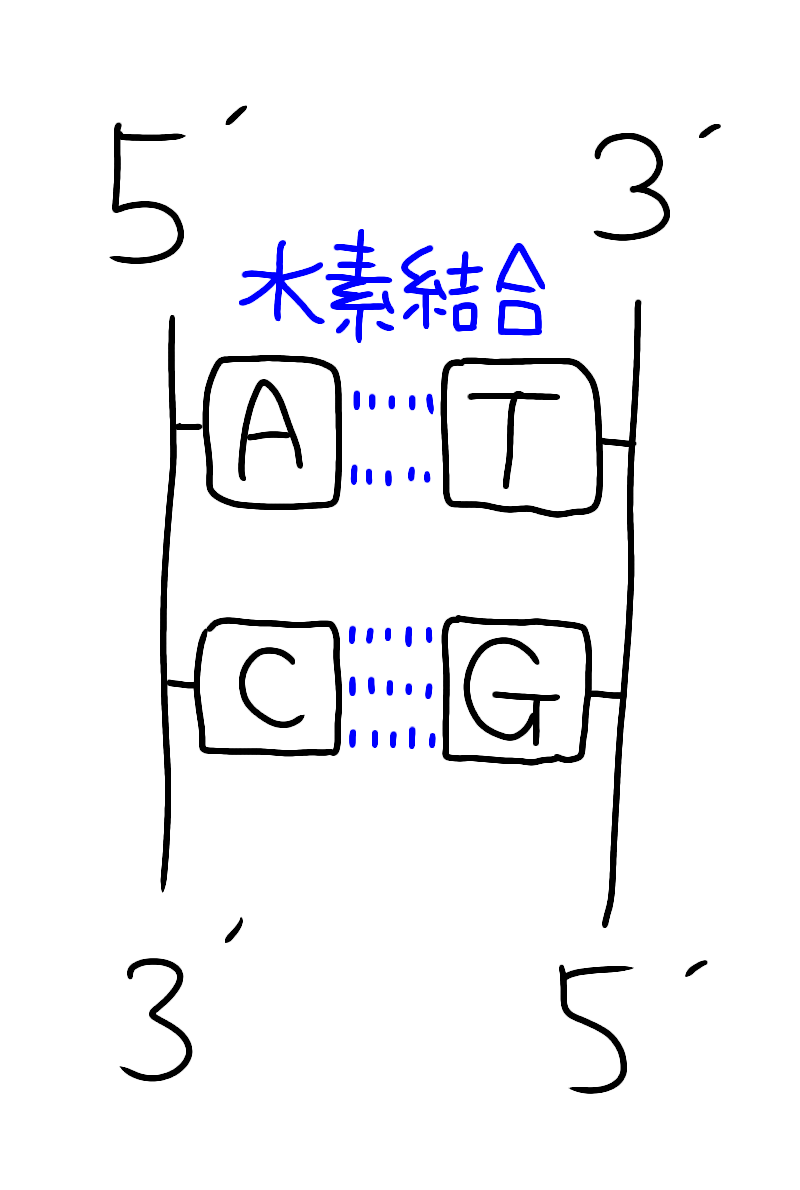
•ذ•û‚جچ½‚ھ3'پ¨5'•ûŒü‚¾‚ئپA‚à‚¤ˆê•û‚ح5'پ¨3'•ûŒü‚ة‚ب‚ء‚ؤ‚¢‚ـ‚·پiپپ‚±‚ê‚ھ‹t•ûŒüگ«پjپB
‚ب‚¨پAƒAƒfƒjƒ“پiAپj‚حƒ`ƒ~ƒ“پiTپj‚ئ‘خ‚ئ‚ب‚èگ…‘fŒ‹چ‡‚ح2–{
ƒVƒgƒVƒ“پiCپj‚حƒOƒAƒjƒ“پiGپj‚ئ‘خ‚ئ‚ب‚èپAگ…‘fŒ‹چ‡‚ح3–{‚إ‚·
پiپ¦CG‚حƒRƒ“ƒsƒ…پ[ƒ^پ[ƒOƒ‰ƒtƒBƒbƒN‚إ3D‚جƒCƒپپ[ƒW‚إ3–{‚ئٹo‚¦‚éپj
•،گ»پE“]ژتپE–|–َ‚ة‚آ‚¢‚ؤ
‚±‚±‚ـ‚إ‚ح–â‘è‚ب‚¢‚ئژv‚¤‚ج‚إ‚·‚ھپA
ˆّ‚ء‚©‚©‚ء‚ؤ‚µ‚ـ‚¤گl‚ھ‘‚¦‚é‚ج‚ھ
پu•،گ»پE“]ژتپE–|–َپv‚جچغ‚جپA•ûŒüگ«‚جکb‚ة‚ب‚ء‚½چغ‚إ‚·پB
پE•،گ»‚حDNA‚©‚çDNA‚ًچ‡گ¬‚·‚锽‰
پE“]ژت‚حDNA‚©‚çRNA‚ًچ‡گ¬‚·‚锽‰
پE–|–َ‚حmRNA‚©‚çƒ^ƒ“ƒpƒNژ؟‚ًچ‡گ¬‚·‚锽‰‚ًژw‚µ‚ـ‚·‚ھ
‚ـ‚ئ‚ك‚é‚ئˆب‰؛‚ج‚و‚¤‚ة‚ب‚è‚ـ‚·
| ’’Œ^‚ئ‚ب‚é‚à‚ج | “ا‚فژو‚ç‚ê‚é’’Œ^‚ج•ûŒü | چ‡گ¬‚³‚ê‚é‚à‚ج | گL’·پiچ‡گ¬پj•ûŒü | |
| •،گ» | DNA | 3'پ¨5'•ûŒü | DNA پiپپƒkƒNƒŒƒIƒ`ƒhپj | 5پfپ¨3'•ûŒü |
| “]ژت | DNA | 3'پ¨5'•ûŒü | RNAپimRNAپj پiپپƒkƒNƒŒƒIƒ`ƒhپj | 5پfپ¨3'•ûŒü |
| –|–َ | mRNAپiپپƒkƒNƒŒƒIƒ`ƒhپj | 5'پ¨3'•ûŒü | ƒ^ƒ“ƒpƒNژ؟ | N––’[پ¨C––’[ |
ڈم‚ج•\‚ج“à—e‚ًٹo‚¦‚éچغ‚ةڈd—v‚ة‚ب‚ء‚ؤ‚‚é•”•ھ‚ًگشگF‚إژ¦‚µ‚ـ‚µ‚½پB
‹ة’[‚بکbپAڈم‚ج•\‚ج“à—e‚ًٹo‚¦‚邽‚ك‚ة‚حگشگF‚ج•”•ھ‚¾‚¯ٹo‚¦‚ê‚خ‚و‚¢‚ج‚إ‚·پB
‚آ‚ـ‚èپuƒkƒNƒŒƒIƒ`ƒh‚جگL’·•ûŒü‚ح5'پ¨3'•ûŒü‚ةŒہ’肳‚ê‚ؤ‚¢‚éپv
ƒkƒNƒŒƒIƒ`ƒh‚حپuƒSƒ~پi5'پ¨3'پj‚ج•ûŒü‚ة‚µ‚©گL‚خ‚¹‚ب‚¢پv‚ئ‚¢‚¤“à—e‚ھچإ‚àڈd—v‚بƒ|ƒCƒ“ƒg‚ة‚ب‚è‚ـ‚·پB
‚±‚ê‚ًٹغˆأ‹L‚·‚é‚ج‚إ‚ح‚ب‚پAچ\‘¢ƒŒƒxƒ‹‚إ—‰ً‚µ‚ـ‚µ‚ه‚¤پB
‰؛‚جƒCƒ‰ƒXƒg‚حƒkƒNƒŒƒIƒ`ƒh‚ھDNAƒ|ƒٹƒپƒ‰پ[ƒ[‚ب‚ا‚إگL’·‚µ‚ؤ‚¢‚‚ج‚ً–حژ®‰»‚µ‚½‚à‚ج‚ة‚ب‚è‚ـ‚·پB
DNA‚ھ5'پ¨3'•ûŒü‚ةگL’·‚·‚éچغ‚ج‰»ٹw”½‰پi–حژ®گ}پj
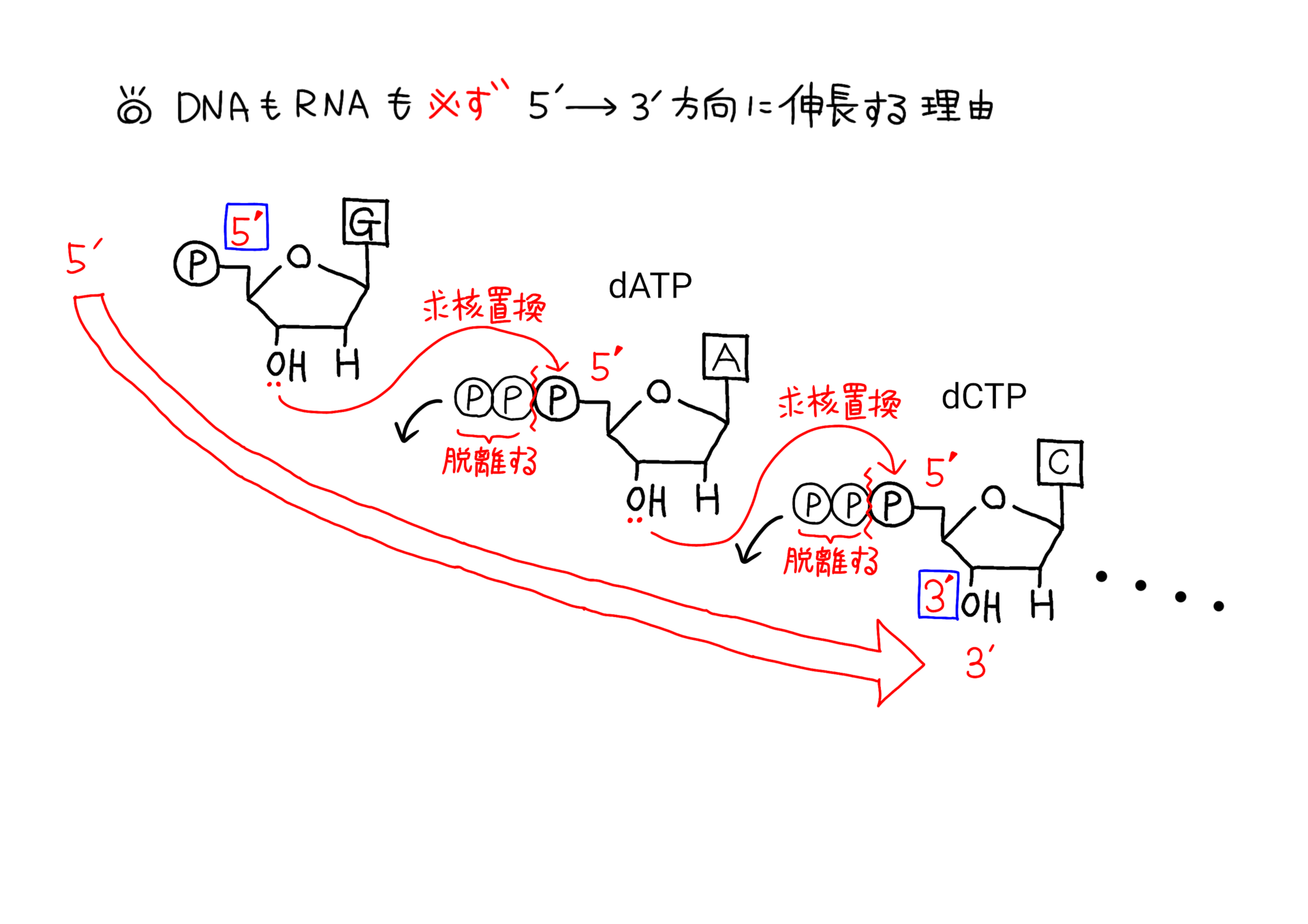
‚±‚جƒCƒ‰ƒXƒg‚ً‚و‚Œ©‚ؤ‚à‚炦‚خپADNAپi‚âRNA‚ئ‚¢‚ء‚½ƒkƒNƒŒƒIƒ`ƒhپj‚ھ5'پ¨3'•ûŒü‚ة‚µ‚©گL’·‚µ‚ب‚¢——R‚ھ‚و‚•ھ‚©‚é‚©‚ئژv‚¢‚ـ‚·پB
‚»‚µ‚ؤپA–`“ھ‚ةگ\‚µڈم‚°‚½‚ئ‚¨‚èپADNAپiƒkƒNƒŒƒIƒ`ƒhپj‚ح2–{چ½‚ًŒ`گ¬‚·‚éچغ‚ة‚ح‹t•ûŒüگ«‚ً‚ئ‚邽‚كپA’’Œ^پi“ا‚فژو‚ç‚ê‚鑤‚جچ½پj‚©‚ç‚جژ‹“_‚إ‚ح•،گ»‚ئ“]ژت‚ة‚¨‚¢‚ؤپA’’Œ^‚حپu3'پ¨5'پv•ûŒü‚ة“ا‚فژو‚ç‚ê‚ؤ‚¢‚‚±‚ئ‚ة‚ب‚éپB
•،گ»‚¾‚¯‚ح•ûŒüگ«‚ة’چˆس
‚±‚±‚ـ‚إ‚ج“à—e‚إپA‚±‚ج•\‚ج•،گ»پE“]ژت‚جچs‚ة‚آ‚¢‚ؤ‚حگà–¾‚ھ‚إ‚«‚ـ‚µ‚½پB
| ’’Œ^‚ئ‚ب‚é‚à‚ج | “ا‚فژو‚ç‚ê‚é’’Œ^‚ج•ûŒü | چ‡گ¬‚³‚ê‚é‚à‚ج | گL’·پiچ‡گ¬پj•ûŒü | |
| •،گ» | DNA | 3'پ¨5'•ûŒü | DNA پiپپƒkƒNƒŒƒIƒ`ƒhپj | 5پfپ¨3'•ûŒü |
| “]ژت | DNA | 3'پ¨5'•ûŒü | RNAپimRNAپj پiپپƒkƒNƒŒƒIƒ`ƒhپj | 5پfپ¨3'•ûŒü |
| –|–َ | mRNAپiپپƒkƒNƒŒƒIƒ`ƒhپj | 5'پ¨3'•ûŒü | ƒ^ƒ“ƒpƒNژ؟ | N––’[پ¨C––’[ |
‚»‚ê‚إ‚حپA–|–َ‚ة‚آ‚¢‚ؤ‚حˆê‘ج‚ا‚ج‚و‚¤‚ةچl‚¦‚é‚ׂ«‚ب‚ج‚إ‚µ‚ه‚¤‚©پB
‚±‚±‚إ‘هژ–‚ب‚ج‚حپAپu–|–َپپƒkƒNƒŒƒIƒ`ƒh‚ًچى‚锽‰‚إ‚ح‚ب‚¢پv‚ئ‚¢‚¤‚±‚ئ‚إ‚·پB
پw–|–َپx‚ئ‚حmRNA‚ً‚à‚ئ‚ةƒ^ƒ“ƒpƒNژ؟‚ًچى‚锽‰‚ب‚ج‚إپA
’’Œ^‚ئ‚ب‚émRNA‚ح—ک_ڈمپA5'پ¨3'•ûŒü‚ة“ا‚فژو‚邱‚ئ‚ة‰½‚çژxڈل‚ح‚ب‚¢‚ج‚إ‚·پB
پiگ®—‚µ‚ؤ‚ظ‚µ‚¢‚ج‚حپAپw•،گ»پx‚ئپw–|–َپx‚حپA‚»‚ꂼ‚êDNA‚âRNA‚ئ‚¢‚¤ƒkƒNƒŒƒIƒ`ƒh‚ًچ‡گ¬‚·‚锽‰‚إ‚ ‚èپA
پ@ƒkƒNƒŒƒIƒ`ƒh‚جگL’·پiچ‡گ¬پj•ûŒü‚ة‚ح5'پ¨3'‚ج‚ف‚ئ‚¢‚¤”›‚è‚ھ‚ ‚èپA•K‘R“I‚ة’’Œ^‚ح3'پ¨5'‚ة“ا‚فژو‚ç‚ê‚é•K—v‚ھ‚ ‚ء‚½‚±‚ئپB–|–َ‚حƒ^ƒ“ƒpƒNژ؟‚ئ‚¢‚¤ƒ|ƒٹƒyƒvƒ`ƒh‚ًچى‚锽‰‚إ‚ ‚èپAN––’[پAC––’[‚ئ‚¢‚¤ٹT”O‚ح‚ ‚é‚ھپA3'‚ئ‚©5'‚ب‚ا‚جٹT”O‚ح•K—v‚ب‚¢‚ئ‚¢‚¤“_‚إ‚ ‚éپj
پw“]ژتپx‚جچغ‚ةmRNA‚ًچ‡گ¬پimRNA‚جƒvƒچƒZƒVƒ“ƒO‚ًپj‚·‚éچغپA
mRNA‚ج5'––’[‚ة‚ح7-ƒپƒ`ƒ‹ƒOƒAƒmƒVƒ“‚ھ•t‰ء‚³‚êپicapچ\‘¢‚ئŒؤ‚شپjپA3'––’[‚ة‚حƒ|ƒٹپiAپjƒeƒCƒ‹‚ئŒؤ‚خ‚ê‚éƒAƒfƒjƒ‹ژ_‚ھ150پ`200Œآ‚ظ‚اکAŒ‹‚µ‚½چ\‘¢‚ھ•t‰ء‚³‚ê‚éپB
‚»‚µ‚ؤmRNA‚حپA5'––’[‚جcapچ\‘¢‚©‚çƒٹƒ{ƒ\پ[ƒ€‚جڈ¬ƒTƒuƒ†ƒjƒbƒgپiگ^ٹjگ¶•¨‚إ‚ح40SپAŒ´ٹjگ¶•¨‚إ‚ح30SƒTƒuƒ†ƒjƒbƒgپj
‚ة‚و‚è“ا‚فژو‚ç‚ê‚ؤ3'––’[‚جƒ|ƒٹپiAپjƒeƒCƒ‹‚ج‚ظ‚¤‚ةŒü‚©‚ء‚ؤ“ا‚فژو‚ç‚ê‚ؤ‚¢‚پB
پ¦5'––’[‚جƒLƒƒƒbƒvچ\‘¢‚ج‹ك‚‚ة‘¶چف‚·‚éٹJژnƒRƒhƒ“‚©‚ç“ا‚فژو‚è‚ًƒXƒ^پ[ƒg‚µ‚ؤپA3'––’[‚جƒ|ƒٹپiAپjƒeƒCƒ‹‘¤‚ةڈIژ~ƒRƒhƒ“‚ھ‘¶چف‚·‚邽‚كپAپumRNA‚حƒLƒƒƒbƒv‚ً”ي‚ء‚ؤ‚¢‚é“ھپi5'پj‚©‚ç“ا‚فژو‚è‚ًٹJژn‚·‚éپv‚ئٹo‚¦‚é
‚µ‚½‚ھ‚ء‚ؤپAŒ‹ک_‚ئ‚µ‚ؤ–|–َ‚ة‚¨‚¢‚ؤ‚ح
’’Œ^mRNA‚ح5'پ¨3'•ûŒü‚ة“ا‚فژو‚ç‚ê‚ؤپA
ƒ^ƒ“ƒpƒNژ؟‚حN––’[پ¨C––’[•ûŒü‚ةگL’·‚ھگi‚ٌ‚إ‚¢‚‚±‚ئ‚ئ‚ب‚éپB
“ٹچeژزپFƒhƒNƒ^پ[PGپb11:36
پb•ھژqگ¶•¨ٹw




