2019年03月04日
DNAの伸長方向は5'→3'方向になる理由を構造レベルで理解しよう
今回は分子生物学の内容です。
分子生物学が勉強していると必ず出てくるDNA鎖の伸長方向についての考え方です。
まず、基礎の内容としてDNAというのは逆方向性の2本鎖ヌクレオチド構造を取りますね。
イメージでいうと以下のような構造です。
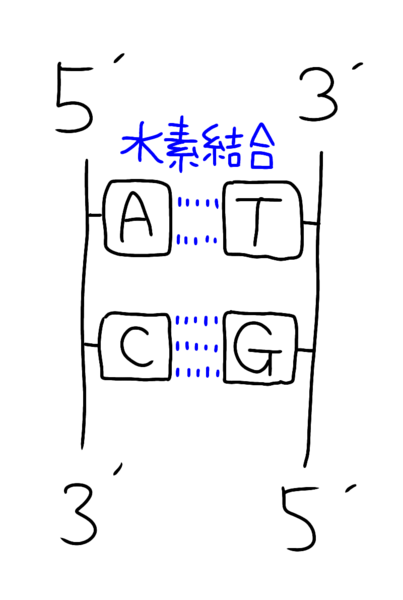
片方の鎖が3'→5'方向だと、もう一方は5'→3'方向になっています(=これが逆方向性)。
なお、アデニン(A)はチミン(T)と対となり水素結合は2本
シトシン(C)はグアニン(G)と対となり、水素結合は3本です
(※CGはコンピューターグラフィックで3Dのイメージで3本と覚える)
ここまでは問題ないと思うのですが、
引っかかってしまう人が増えるのが
「複製・転写・翻訳」の際の、方向性の話になった際です。
・複製はDNAからDNAを合成する反応
・転写はDNAからRNAを合成する反応
・翻訳はmRNAからタンパク質を合成する反応を指しますが
まとめると以下のようになります
上の表の内容を覚える際に重要になってくる部分を赤色で示しました。
極端な話、上の表の内容を覚えるためには赤色の部分だけ覚えればよいのです。
つまり「ヌクレオチドの伸長方向は5'→3'方向に限定されている」
ヌクレオチドは「ゴミ(5'→3')の方向にしか伸ばせない」という内容が最も重要なポイントになります。
これを丸暗記するのではなく、構造レベルで理解しましょう。
下のイラストはヌクレオチドがDNAポリメラーゼなどで伸長していくのを模式化したものになります。
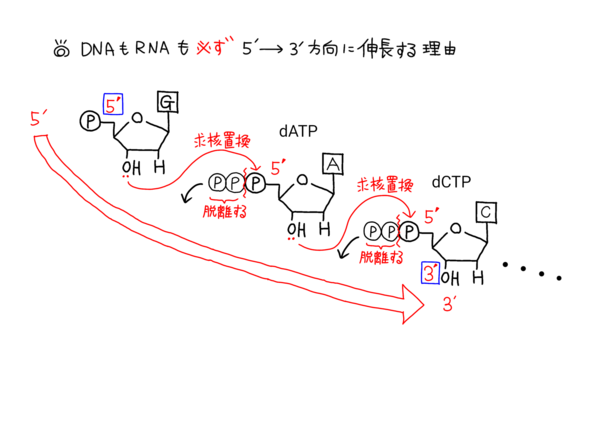
このイラストをよく見てもらえば、DNA(やRNAといったヌクレオチド)が5'→3'方向にしか伸長しない理由がよく分かるかと思います。
そして、冒頭に申し上げたとおり、DNA(ヌクレオチド)は2本鎖を形成する際には逆方向性をとるため、鋳型(読み取られる側の鎖)からの視点では複製と転写において、鋳型は「3'→5'」方向に読み取られていくことになる。
ここまでの内容で、この表の複製・転写の行については説明ができました。
それでは、翻訳については一体どのように考えるべきなのでしょうか。
ここで大事なのは、「翻訳=ヌクレオチドを作る反応ではない」ということです。
『翻訳』とはmRNAをもとにタンパク質を作る反応なので、
鋳型となるmRNAは理論上、5'→3'方向に読み取ることに何ら支障はないのです。
(整理してほしいのは、『複製』と『翻訳』は、それぞれDNAやRNAというヌクレオチドを合成する反応であり、
ヌクレオチドの伸長(合成)方向には5'→3'のみという縛りがあり、必然的に鋳型は3'→5'に読み取られる必要があったこと。翻訳はタンパク質というポリペプチドを作る反応であり、N末端、C末端という概念はあるが、3'とか5'などの概念は必要ないという点である)
『転写』の際にmRNAを合成(mRNAのプロセシングを)する際、
mRNAの5'末端には7-メチルグアノシンが付加され(cap構造と呼ぶ)、3'末端にはポリ(A)テイルと呼ばれるアデニル酸が150~200個ほど連結した構造が付加される。
そしてmRNAは、5'末端のcap構造からリボソームの小サブユニット(真核生物では40S、原核生物では30Sサブユニット)
により読み取られて3'末端のポリ(A)テイルのほうに向かって読み取られていく。
※5'末端のキャップ構造の近くに存在する開始コドンから読み取りをスタートして、3'末端のポリ(A)テイル側に終止コドンが存在するため、「mRNAはキャップを被っている頭(5')から読み取りを開始する」と覚える
したがって、結論として翻訳においては
鋳型mRNAは5'→3'方向に読み取られて、
タンパク質はN末端→C末端方向に伸長が進んでいくこととなる。
分子生物学が勉強していると必ず出てくるDNA鎖の伸長方向についての考え方です。
まず、基礎の内容としてDNAというのは逆方向性の2本鎖ヌクレオチド構造を取りますね。
イメージでいうと以下のような構造です。
DNAの逆方向性
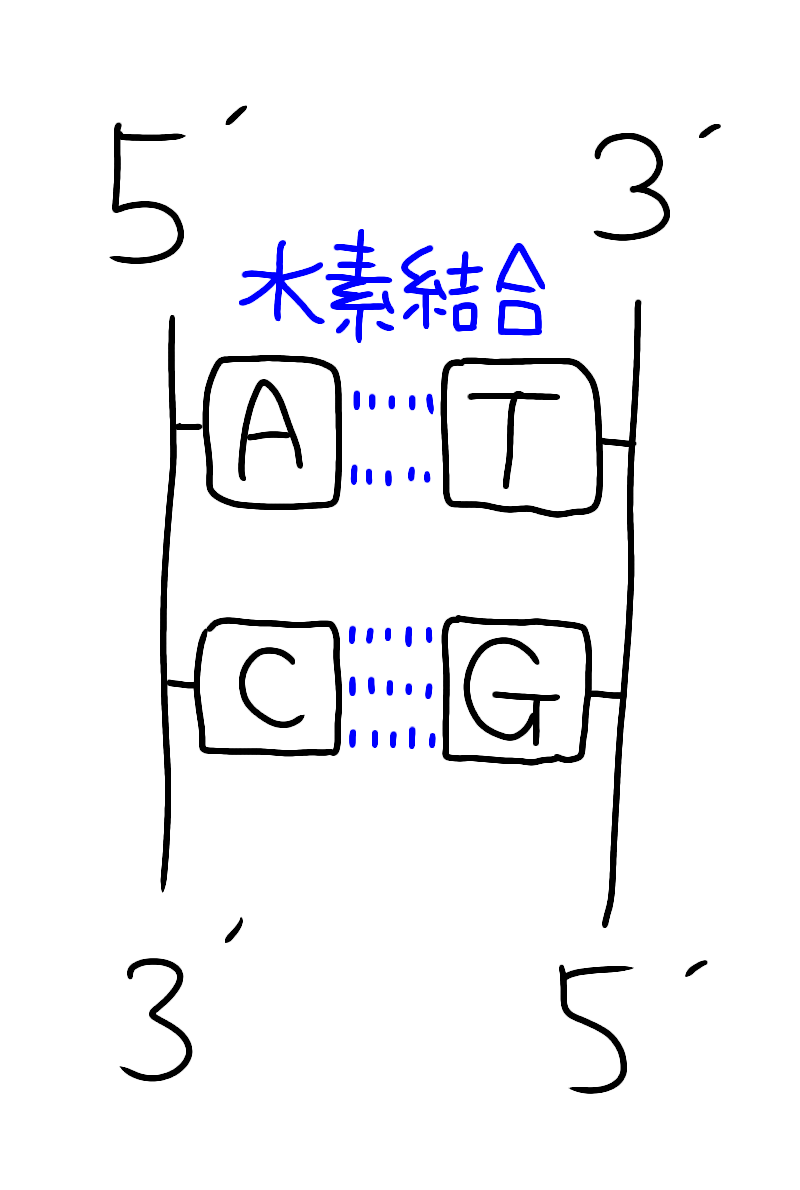
片方の鎖が3'→5'方向だと、もう一方は5'→3'方向になっています(=これが逆方向性)。
なお、アデニン(A)はチミン(T)と対となり水素結合は2本
シトシン(C)はグアニン(G)と対となり、水素結合は3本です
(※CGはコンピューターグラフィックで3Dのイメージで3本と覚える)
複製・転写・翻訳について
ここまでは問題ないと思うのですが、
引っかかってしまう人が増えるのが
「複製・転写・翻訳」の際の、方向性の話になった際です。
・複製はDNAからDNAを合成する反応
・転写はDNAからRNAを合成する反応
・翻訳はmRNAからタンパク質を合成する反応を指しますが
まとめると以下のようになります
| 鋳型となるもの | 読み取られる鋳型の方向 | 合成されるもの | 伸長(合成)方向 | |
| 複製 | DNA | 3'→5'方向 | DNA (=ヌクレオチド) | 5’→3'方向 |
| 転写 | DNA | 3'→5'方向 | RNA(mRNA) (=ヌクレオチド) | 5’→3'方向 |
| 翻訳 | mRNA(=ヌクレオチド) | 5'→3'方向 | タンパク質 | N末端→C末端 |
上の表の内容を覚える際に重要になってくる部分を赤色で示しました。
極端な話、上の表の内容を覚えるためには赤色の部分だけ覚えればよいのです。
つまり「ヌクレオチドの伸長方向は5'→3'方向に限定されている」
ヌクレオチドは「ゴミ(5'→3')の方向にしか伸ばせない」という内容が最も重要なポイントになります。
これを丸暗記するのではなく、構造レベルで理解しましょう。
下のイラストはヌクレオチドがDNAポリメラーゼなどで伸長していくのを模式化したものになります。
DNAが5'→3'方向に伸長する際の化学反応(模式図)
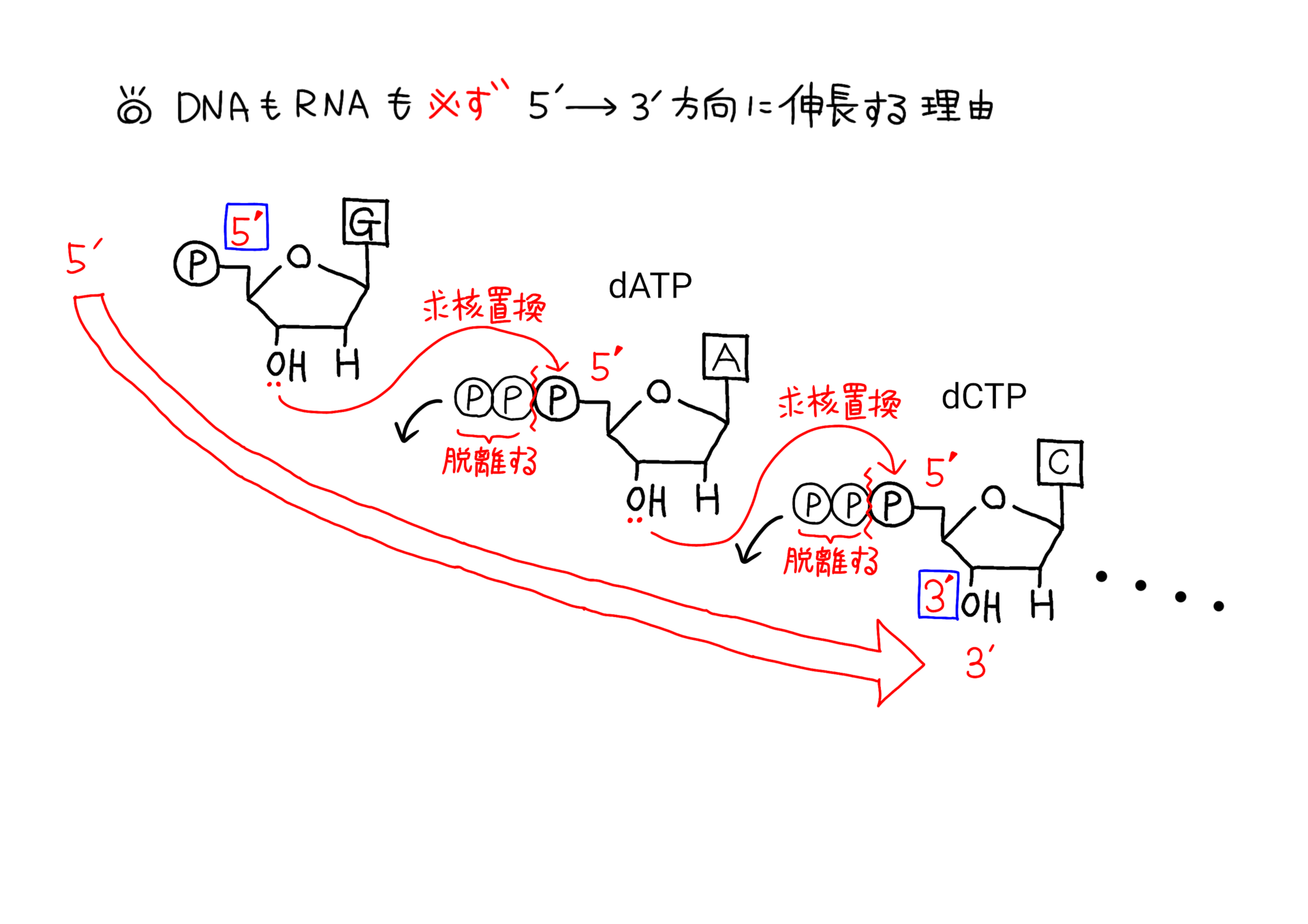
このイラストをよく見てもらえば、DNA(やRNAといったヌクレオチド)が5'→3'方向にしか伸長しない理由がよく分かるかと思います。
そして、冒頭に申し上げたとおり、DNA(ヌクレオチド)は2本鎖を形成する際には逆方向性をとるため、鋳型(読み取られる側の鎖)からの視点では複製と転写において、鋳型は「3'→5'」方向に読み取られていくことになる。
複製だけは方向性に注意
ここまでの内容で、この表の複製・転写の行については説明ができました。
| 鋳型となるもの | 読み取られる鋳型の方向 | 合成されるもの | 伸長(合成)方向 | |
| 複製 | DNA | 3'→5'方向 | DNA (=ヌクレオチド) | 5’→3'方向 |
| 転写 | DNA | 3'→5'方向 | RNA(mRNA) (=ヌクレオチド) | 5’→3'方向 |
| 翻訳 | mRNA(=ヌクレオチド) | 5'→3'方向 | タンパク質 | N末端→C末端 |
それでは、翻訳については一体どのように考えるべきなのでしょうか。
ここで大事なのは、「翻訳=ヌクレオチドを作る反応ではない」ということです。
『翻訳』とはmRNAをもとにタンパク質を作る反応なので、
鋳型となるmRNAは理論上、5'→3'方向に読み取ることに何ら支障はないのです。
(整理してほしいのは、『複製』と『翻訳』は、それぞれDNAやRNAというヌクレオチドを合成する反応であり、
ヌクレオチドの伸長(合成)方向には5'→3'のみという縛りがあり、必然的に鋳型は3'→5'に読み取られる必要があったこと。翻訳はタンパク質というポリペプチドを作る反応であり、N末端、C末端という概念はあるが、3'とか5'などの概念は必要ないという点である)
『転写』の際にmRNAを合成(mRNAのプロセシングを)する際、
mRNAの5'末端には7-メチルグアノシンが付加され(cap構造と呼ぶ)、3'末端にはポリ(A)テイルと呼ばれるアデニル酸が150~200個ほど連結した構造が付加される。
そしてmRNAは、5'末端のcap構造からリボソームの小サブユニット(真核生物では40S、原核生物では30Sサブユニット)
により読み取られて3'末端のポリ(A)テイルのほうに向かって読み取られていく。
※5'末端のキャップ構造の近くに存在する開始コドンから読み取りをスタートして、3'末端のポリ(A)テイル側に終止コドンが存在するため、「mRNAはキャップを被っている頭(5')から読み取りを開始する」と覚える
したがって、結論として翻訳においては
鋳型mRNAは5'→3'方向に読み取られて、
タンパク質はN末端→C末端方向に伸長が進んでいくこととなる。
【分子生物学の最新記事】



この記事へのコメント