2019年05月14日
基底核の機能構造
最終的には、心の働きの脳内メカニズムについて述べていきます。
基底核の機能構造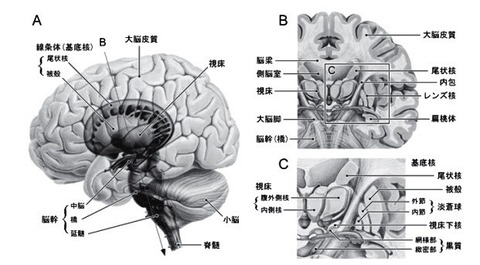
2―1;基底核の構築(図 2)
基底核は大脳半球の深部に存在する.
基底核は内包により尾状核とレンズ核(被殻・淡蒼球)に分けられ,内側には視床が存在する.
尾状核の尾部は辺縁系の一部である扁桃体へと連続する.
基底核は,
(新)線条体(Neostriatum),
淡蒼球(Globus pallidus),
黒質(Substantia nigra),
視床下核(Subthalamic nucleus)の4領域に分られる.
線条体は尾状核(Caudate nucleus)と被殻(Putamen)とからなり,
淡蒼球は,内節(Internal segment;GPi)と外節(Externalsegment;GPe)とからなる.
黒質(Substantianigra)は,緻密部(pars compacta;SNc)と網様部(pars reticulata;SNr)とに分けられる.
線条体は大脳皮質の広範な領域から入力を受ける.
淡蒼球内節と黒質網様部は基底核の出力核である.
その出力はγ-アミノ酪酸(gammaaminobutylic acid;GABA)を
神経伝達物質とする抑制性投射である.
視床下核,淡蒼球外節,黒質緻密部は基底核内の各々の神経核と線維連絡を持ち,
基底核内の神経回路活動を修飾する.
黒質緻密部は,
腹側被蓋野(Ventral tegmental nucleus;VTA)や
後赤核領域(Retrorubral field)と共に
神経伝達物質としてドーパミンを有している.
2―2;線条体の微細構造と神経細胞群
線条体は均一の構造に見えるが,マトリックス(Matrix)とパッチ(Patch)という2つのコンパートメントに分けられ,各々の領域は次の様な入出力系の特徴を持っている.
①マトリックスは主に大脳皮質のIII層とVa層から,パッチはVb層と VI 層からの投射を受ける.
②新皮質はパッチよりもマトリックスに多く投射し,その他の皮質はパッチに多く投射する.
③マトリックスに分布する投射ニューロンは淡蒼球や黒質網様部に投射し,
パッチに分布する投射ニューロンは主に黒質緻密部に投射する[5― 8].
線条体の80~95%の細胞は線条体外へ線維を送る投射ニューロンであり,
GABAを伝達物質とする中型の有棘細胞(Medium spiny neurons)である.
5~20%の細胞は線条体内の介在細胞である.
マウスやラットに比べて霊長類(サル)では投射細胞の比率は低い[9].
投射細胞は次の2群に大別される.
一群はGABAとエンケファリン(Enkephalin)やニューロテンシン(Neurotensin)を含有し,
ドーパミンD2受容体を持つ.
これらのニューロンは淡蒼球外節に投射して間接路を構成する.
もう一群の細胞はGABAとサブスタンスP(Substance P)やダイノルフィン(Dynorphine)を含有し,
ドーパミンD1受容体を有する.
そして淡蒼球内節や黒質網様部に投射して直接路を構成する[10, 11].
介在細胞には,
①大型のコリン作動性ニューロン,
②中型でソマトスタチン(Somatostatin),ニューロペプチドY(Neuropeptide Y),一酸化窒素(Nitric oxide)を持つニューロン,
③中型で GABAを伝達物質とし,カルシウム結合蛋白質であるパルブアルブミン(Parvalbumin)を含
有するニューロンなどが存在する[10].
中でもコリン作動性ニューロンについての解析が進んでいる.
中脳からのドーパミン作動性投射とコリン作動性ニューロンとの相互作用が投射ニューロンの活動を調節する上で重要であり,基底核疾患ではこの相互作用がしばしば破綻する[11― 13].
【引用文献】
大脳基底核の機能;パーキンソン病との関連において
旭川医科大学 生理学第二講座
高草木 薫
参考文献
5.Gerfen CR : The neostriatal mosaic : striatal patchmatrix organization is related to cortical lamination.
Science 246 : 385― 388, 1989.
6.Gerfen CR : The naostriatal mosaic : multiple levels of
compartmental organization in the basal ganglia.
Annu Rev Neurosci 15 : 285― 320, 1992.
7.Graybiel AM : Neurotransmitters and neuromodulators in the basal ganglia. Trends Neurosci 13 : 244―
254, 1990.
8.Kincaid AE & Wilson CJ : Corticostriatal innervation
of the patch and matrix in the rat neostriatum. J
Comp Neurol 374 : 578― 592, 1996.
9.Graveland GA & Difiglia M : The frequency and distribution of medium-sized neurons with indented nuclei in the primate and rodent neostriatum. Brain Res
327, 307― 311, 1985.
10.Kawaguchi Y : Neostriatal cell subtypes and their
functional roles. Neurosci Res 27 : 1― 8, 1997.
11.Mitchell IJ, Cooper AJ & Griffiths MR : The selective
vulnerability of striatopallidal neurons. Prog Neurobiol 59 : 691― 719, 1999.
12.Cicchetti F, Prensa L, Wu Y & Parent A : 2000.
Chemical anatomy of striatum interneurons in normal and individuals and in patients with Huntington’s
disease. Brain Res Rev 34 : 80― 101.
13.Zou F-M, Wilson CJ & Dani JA : Cholinergic interneuron characteristics and nicotinic properties in
the striatum. J Neurobiol 53 : 590― 605, 2002.
基底核の機能構造
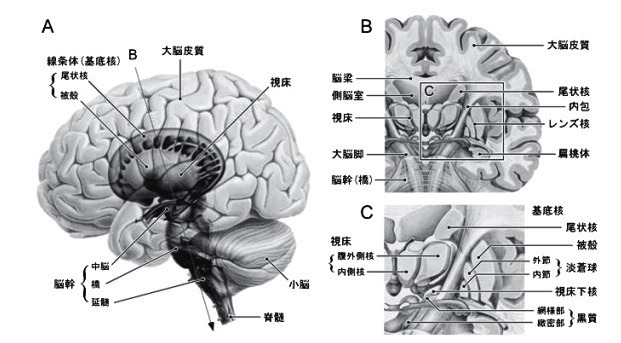
2―1;基底核の構築(図 2)
基底核は大脳半球の深部に存在する.
基底核は内包により尾状核とレンズ核(被殻・淡蒼球)に分けられ,内側には視床が存在する.
尾状核の尾部は辺縁系の一部である扁桃体へと連続する.
基底核は,
(新)線条体(Neostriatum),
淡蒼球(Globus pallidus),
黒質(Substantia nigra),
視床下核(Subthalamic nucleus)の4領域に分られる.
線条体は尾状核(Caudate nucleus)と被殻(Putamen)とからなり,
淡蒼球は,内節(Internal segment;GPi)と外節(Externalsegment;GPe)とからなる.
黒質(Substantianigra)は,緻密部(pars compacta;SNc)と網様部(pars reticulata;SNr)とに分けられる.
線条体は大脳皮質の広範な領域から入力を受ける.
淡蒼球内節と黒質網様部は基底核の出力核である.
その出力はγ-アミノ酪酸(gammaaminobutylic acid;GABA)を
神経伝達物質とする抑制性投射である.
視床下核,淡蒼球外節,黒質緻密部は基底核内の各々の神経核と線維連絡を持ち,
基底核内の神経回路活動を修飾する.
黒質緻密部は,
腹側被蓋野(Ventral tegmental nucleus;VTA)や
後赤核領域(Retrorubral field)と共に
神経伝達物質としてドーパミンを有している.
2―2;線条体の微細構造と神経細胞群
線条体は均一の構造に見えるが,マトリックス(Matrix)とパッチ(Patch)という2つのコンパートメントに分けられ,各々の領域は次の様な入出力系の特徴を持っている.
①マトリックスは主に大脳皮質のIII層とVa層から,パッチはVb層と VI 層からの投射を受ける.
②新皮質はパッチよりもマトリックスに多く投射し,その他の皮質はパッチに多く投射する.
③マトリックスに分布する投射ニューロンは淡蒼球や黒質網様部に投射し,
パッチに分布する投射ニューロンは主に黒質緻密部に投射する[5― 8].
線条体の80~95%の細胞は線条体外へ線維を送る投射ニューロンであり,
GABAを伝達物質とする中型の有棘細胞(Medium spiny neurons)である.
5~20%の細胞は線条体内の介在細胞である.
マウスやラットに比べて霊長類(サル)では投射細胞の比率は低い[9].
投射細胞は次の2群に大別される.
一群はGABAとエンケファリン(Enkephalin)やニューロテンシン(Neurotensin)を含有し,
ドーパミンD2受容体を持つ.
これらのニューロンは淡蒼球外節に投射して間接路を構成する.
もう一群の細胞はGABAとサブスタンスP(Substance P)やダイノルフィン(Dynorphine)を含有し,
ドーパミンD1受容体を有する.
そして淡蒼球内節や黒質網様部に投射して直接路を構成する[10, 11].
介在細胞には,
①大型のコリン作動性ニューロン,
②中型でソマトスタチン(Somatostatin),ニューロペプチドY(Neuropeptide Y),一酸化窒素(Nitric oxide)を持つニューロン,
③中型で GABAを伝達物質とし,カルシウム結合蛋白質であるパルブアルブミン(Parvalbumin)を含
有するニューロンなどが存在する[10].
中でもコリン作動性ニューロンについての解析が進んでいる.
中脳からのドーパミン作動性投射とコリン作動性ニューロンとの相互作用が投射ニューロンの活動を調節する上で重要であり,基底核疾患ではこの相互作用がしばしば破綻する[11― 13].
【引用文献】
大脳基底核の機能;パーキンソン病との関連において
旭川医科大学 生理学第二講座
高草木 薫
参考文献
5.Gerfen CR : The neostriatal mosaic : striatal patchmatrix organization is related to cortical lamination.
Science 246 : 385― 388, 1989.
6.Gerfen CR : The naostriatal mosaic : multiple levels of
compartmental organization in the basal ganglia.
Annu Rev Neurosci 15 : 285― 320, 1992.
7.Graybiel AM : Neurotransmitters and neuromodulators in the basal ganglia. Trends Neurosci 13 : 244―
254, 1990.
8.Kincaid AE & Wilson CJ : Corticostriatal innervation
of the patch and matrix in the rat neostriatum. J
Comp Neurol 374 : 578― 592, 1996.
9.Graveland GA & Difiglia M : The frequency and distribution of medium-sized neurons with indented nuclei in the primate and rodent neostriatum. Brain Res
327, 307― 311, 1985.
10.Kawaguchi Y : Neostriatal cell subtypes and their
functional roles. Neurosci Res 27 : 1― 8, 1997.
11.Mitchell IJ, Cooper AJ & Griffiths MR : The selective
vulnerability of striatopallidal neurons. Prog Neurobiol 59 : 691― 719, 1999.
12.Cicchetti F, Prensa L, Wu Y & Parent A : 2000.
Chemical anatomy of striatum interneurons in normal and individuals and in patients with Huntington’s
disease. Brain Res Rev 34 : 80― 101.
13.Zou F-M, Wilson CJ & Dani JA : Cholinergic interneuron characteristics and nicotinic properties in
the striatum. J Neurobiol 53 : 590― 605, 2002.





この記事へのコメント
コメントを書く
この記事へのトラックバックURL
https://fanblogs.jp/tb/8799858
※ブログオーナーが承認したトラックバックのみ表示されます。
この記事へのトラックバック






